
写在前面
本书是诺贝尔奖获得者埃里克·坎德尔的自传,将个人成长融入科学发展历程娓娓道来,讲述了其传奇的人生探索之旅,书中内容跨越认知心理学、神经科学、细胞生物学多个学科,科普性非常好,亦能从书中领略科研泰斗的治学之道和大家风范。非常不错的一本书!下文是阅读本书时的一些经典摘录。PS:作者领衔主编的《Principles of Neural Science》是国际上最权威神经科学教科书,被称为“神经科学圣经”,豆瓣评分高达9.9分,同样力荐。
序言
认知神经科学是研究人类认知与智能活动的本质及其神经基础的科学,在分子(基因)、细胞、网络(神经回路)、脑区、全脑等各个水平上对人类所有初级和高级的精神活动的心理过程和神经机制,包括感知觉、运动、注意思维情绪、意识等方面开展研究。
认知分子生物学,其核心思想采用分了生物学的知识和方法来探索人类生命的未解之谜。
这一新兴科学基于以下五大基本原理:
第一,脑与心理的合二为一。大脑是一个复杂的具有计算能力的生物器官,它构建我们对世界的感知,调节我们的思维和情绪,并且控制我们的行为,不仅包括那构此简单的运动行为如奔跑和进食,还包括人类特有的复杂行为如思维、言语和艺术创造。这个角度来看,心理是由大脑执行的一系列操作,就像行走是由腿完成的连贯动作一样,不过心理加工要比走路复杂得多。
第二,大脑的每种心理功能,从最简单的反射到最富创造性的语言、音乐和艺术都是由不同脑区的特定神经回路来实现的。这就是为什么我们更倾向于使用“广义的认知神经生物学”,因为它能够反映出心理加工是由一些特定的神经回路来的,而“狭义的认知神经生物学”意味的是一种心理加工的全部操作都只由某个单独的脑区来完成。
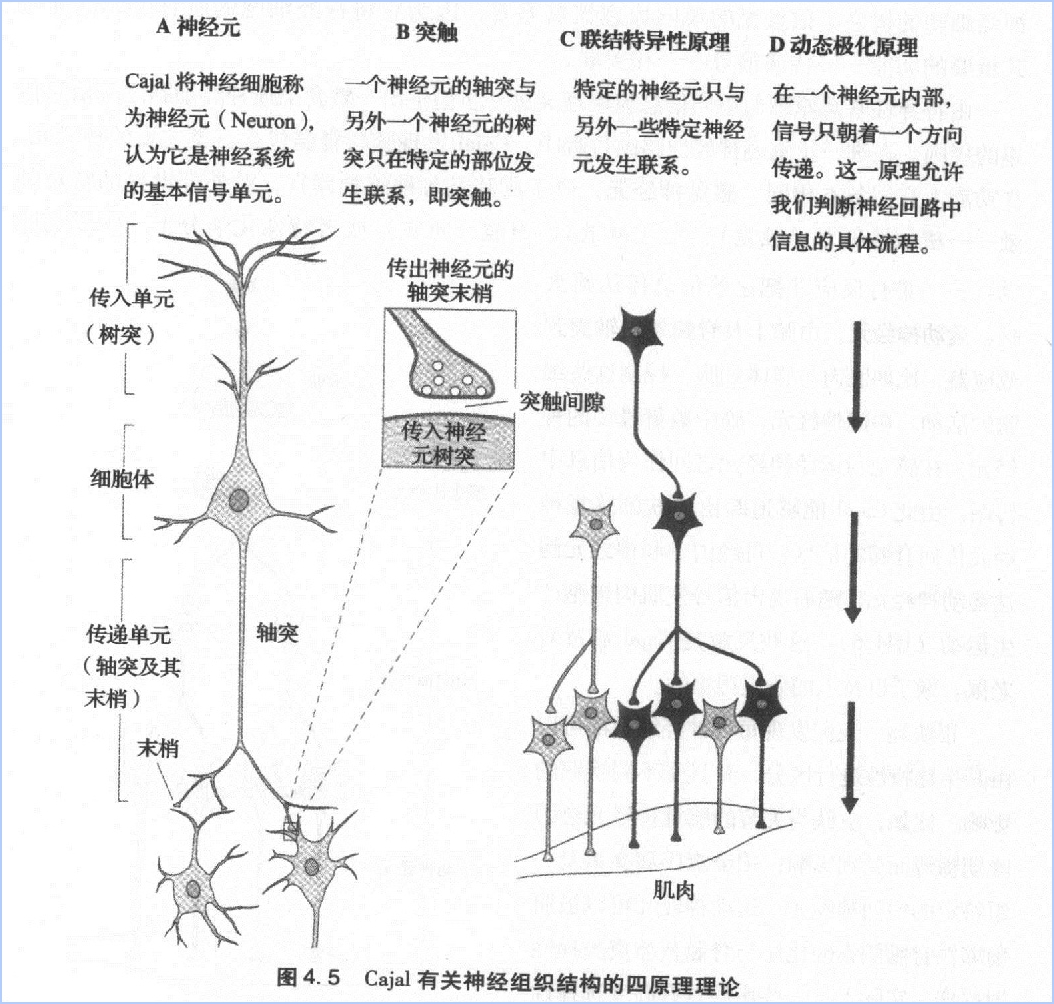
第三,所有这些神经回路都是由相同的基本信号单位—神经元组成的。
第四,神经回路通过细胞内的特殊分子在其内部以及神经细胞之间发出信号。
第五,在数百万年的进化中,这些特殊的信号分子几乎完全被保留了下来。其中有些分子不但存在于我们最远古祖先的细胞中,也存在于现有的进化而来的单细胞生物如细菌、酵母)和简单的多细胞生物(如蠕虫、苍蝇与蜗牛)的细胞中。无论古老还是现代的生物学机制,都是通过分子来控制我们的日常生活,并使得我们适应瞬息万变的环境。
第一卷
除了生物学的存在,影响我们生活的不是真实的过去,而是我们对过去的印象。这些印象常常是结构严密且经过重重筛选的,有如神话般不可言喻。对过去的印象和象征性的构建就像遗传信息一样刻入我们的意识中。每一个历史时期都在当代和过去的神话背景中映射着其本身。——选自 George Steiner,《益胡子蛐城堡》(In Bluebeards Castle,1971)
第二卷
生物学这个领域其实满了无限的可能。我们期待着它给我们带来令人惊叹的消息,人们无法想像将来几十年后的发现又会是什么样子…也许是那时的答案可以将目前人为构造的种种假设理论全部推翻。——弗洛伊德,《超越快乐原则》(Beyond the pleasure principle,1920)
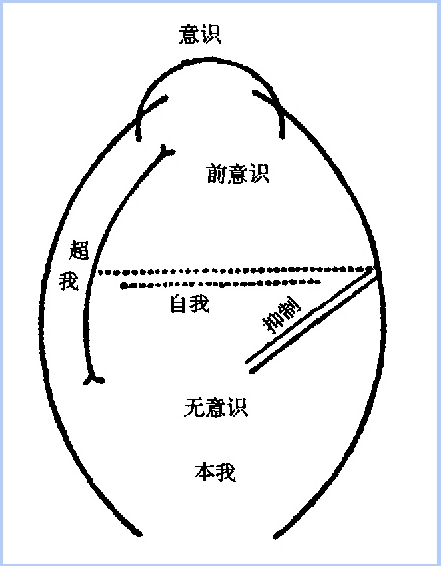 |
弗洛伊德设想人具有三个主要的心理结构:自我、本我以及超我。自我包含有一个意识成分(即有知觉的意识),负责接收感觉信息,并直接与外界相联系;自我同时也包含有一个前意识成分,即一种与意识相连的无意识加工。自我的前意识成分通过压抑与其他防御机制抑制本我的本能冲动,其中包括性与攻击性冲动。自我也同时对于超我的压力进行反应,超我则是一个巨大的无意识的价值载体。虚线表示其所区分的加工是完全无意识的,但又随时与意识相逼。(From NewLectures on Psychoanaly,1933)
Sherrington很快地发现了抑制作用的重要意义,不但协同反射反应,而且能够增强反应稳定性。
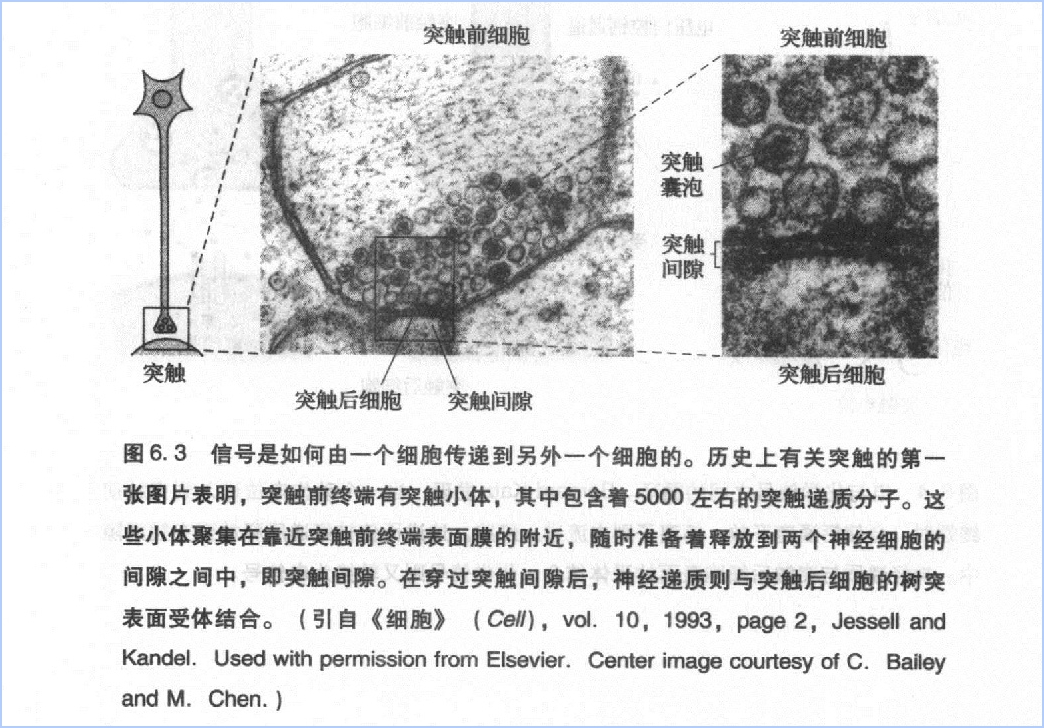
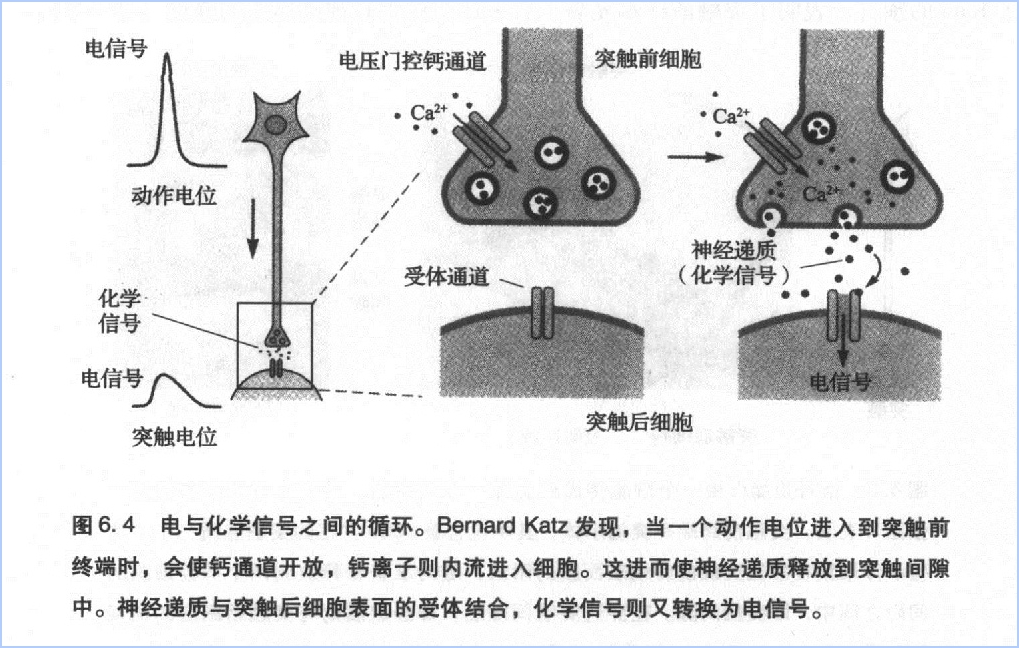
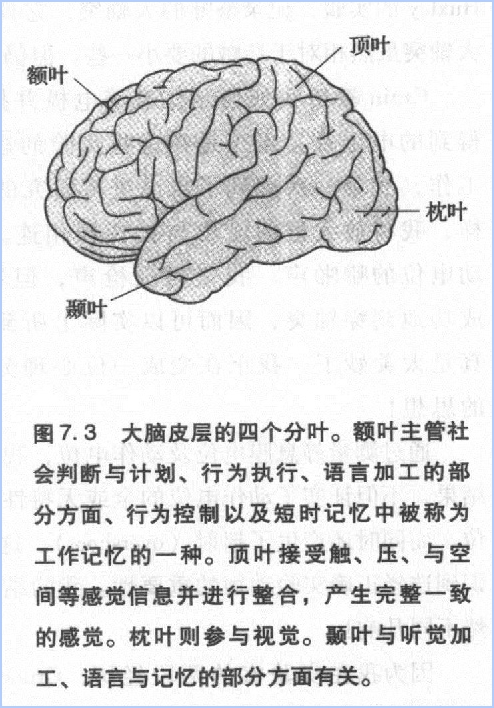
大脑皮层可以分为4个部分:额叶、顶叶、颞叶、枕叶(图7.3)。如果将其展开,人类脑皮质就似一块餐巾布那样大小,只是稍厚一些。它包含上千亿个神经元,每个神经元又都有成千的突触,因此人脑总共约有近10^15个突触结构。
年轻人则应该具有开放的头脑,争取到高手云集的环境中去工作。
大脑皮层两个重要特征。第一,尽管它的两个半球看起来彼此相似,但实际们在结构与功能上都有显著的不同。第二,每个半球分别负责感觉与移动对侧肢体,因此,由左侧肢体(比如说左手)发出到达左侧脊髓的感觉信息最终投射到右侧大脑皮层。同样,右半球运动风其实负责左侧肢体的运动。
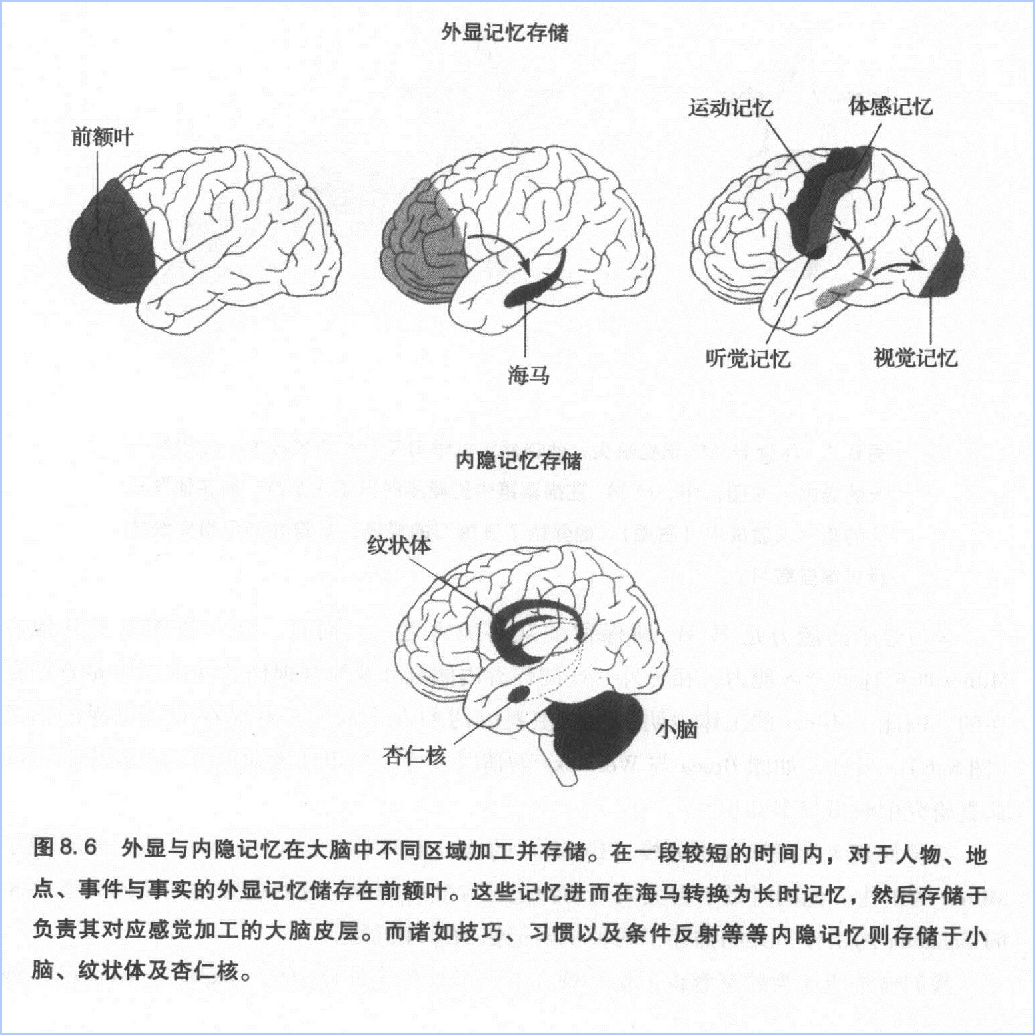
内隐记忆并非是一个单一的记忆系统,它是一组包括大脑皮层下面几个不脑结构的加工集合(图8.6)。例如,与事件相关的感觉联系(害怕或者高兴)就主要做杏仁核的结构有关。形成新的运动或者是认知习惯,与纹状体有关,而学习新的运动技能或者是协调活动则与小脑有关。在包括无脊椎动物的最简单动物中,与习惯、敏感化以及经典条件反射有关的内隐记忆甚至可以储存在反射通路中。
其实人一生中会有很多次类似的情况,即你不可能单单依据冰冷的事实而做出合理的决定—因为事实其实总是不足够的。人最终还是要相信他的潜意识,他的直觉,他的创造性冲动。
第三卷
即将结束的这个世纪是核酸和蛋白的世纪。下个世纪人们将更加关注记忆和欲望。科学家们能够回答自己提出的问题吗?——Francois Jacob,《关于蝇、鼠和人》(Of Flies,Mice and Men,1998)
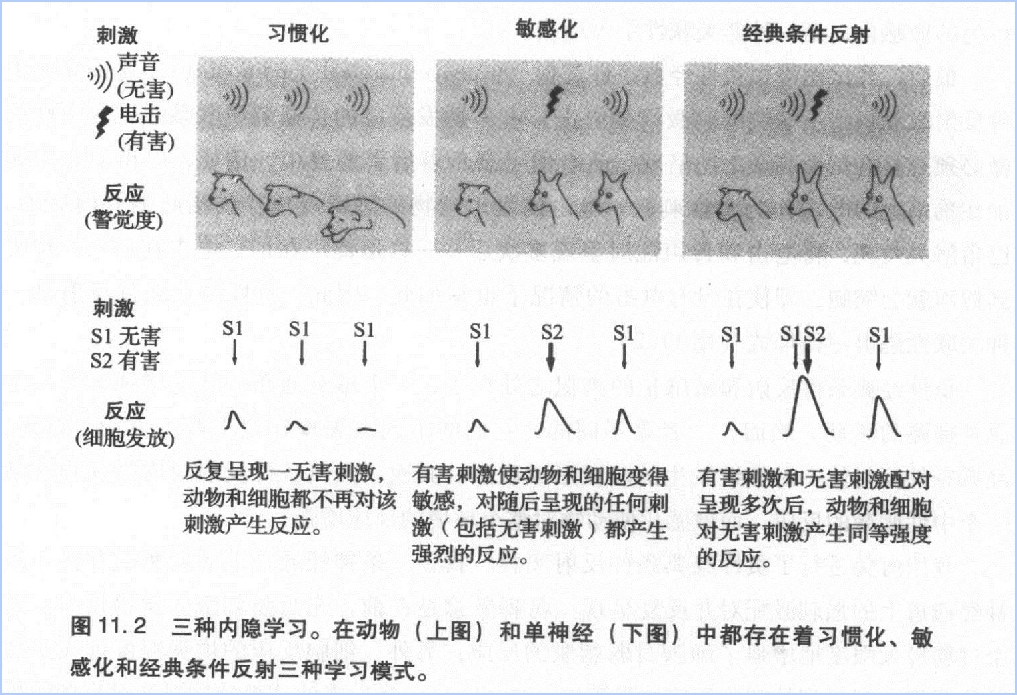
当我决定采用海兔做研究时,我学会了相信自己的直觉,也就是跟着感觉走。成熟的科学家需要具备许多品质,品位可以说是其中最重要的一项了。就像欣赏艺术品音乐、美食和美酒一样,搞科学也要有品位。你要学会分辨重要的和不重要的科学问题。我能感觉到自己品位的提升,我已经学会辨别有趣的问题和无趣的问题——在有趣的事情中,我又能分辨出哪些是可行的。
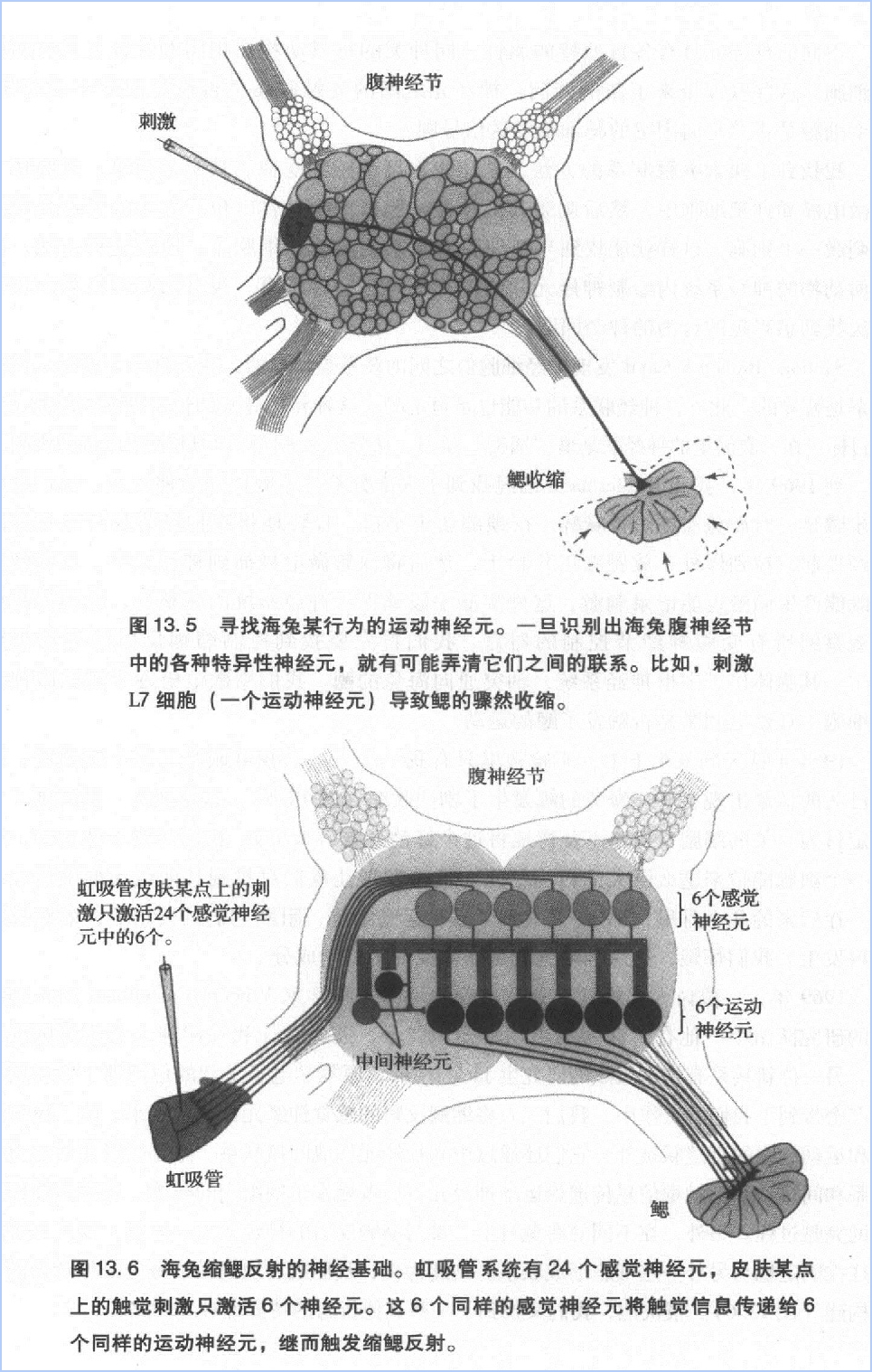
习惯化、敏感化和经典条件反射的实验结果迫使我们思考这样一个问题:经验如何与遗传和发育因素一起影响心理活动的神经基础?遗传和发育决定了神经元之间的连接方式—谁与谁连接,在何时形成连接。然而,遗传和发育因素决定不了突触联系的强度。经验对突触联系强度起到了重要的调节作用。也许遗传和发育因素只能通过先天形成的神经通道控制动物的行为潜能,而环境和学习能够改变神经通道的传递效能,从而带来新的行为形式。总之,我们的海兔研究证明了学习能够改变神经联系强度的观点。
通过还原法研究,我们总结出几条学习和记忆的细胞生物学原则。
首先,突触联系强度的变化足以改变原有的神经网络结构及其信息加工能力。
其次,不同形式的学习可以以完全相反的方式改变两神经元间的突触联系—强化或弱化。
此外,由于缩鰓反射神经回路上的多个位点都发生了变化,所以记忆的储存应该是分布式的,而不是局限在某个点上。
第三,短时记忆存储的时间长短取决于突触强化或弱化的时间长短。
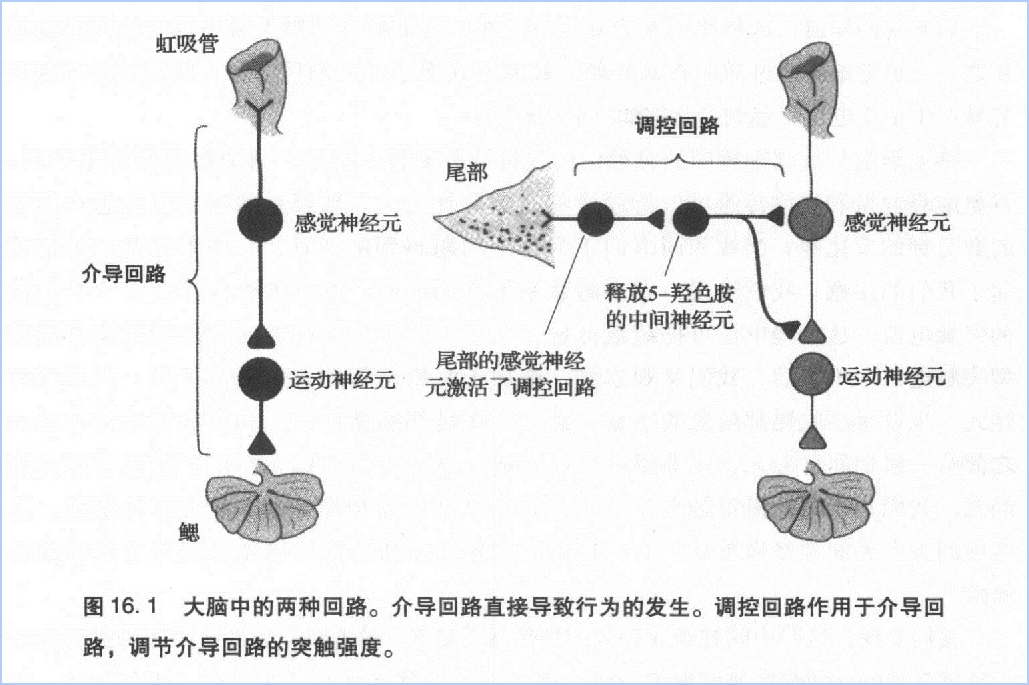
第四,突触强度的调控方式分为同突触和异突触两种。究竟采取哪种方式,取决于学习激活了哪个神经回路——是介导回路(mediating circuit),还是调制回路(modulatory circuit)。对海兔的缩鳃反射前言,介导回路由虹吸管的感觉神经元、中间神经元运动神经元构成。调制回路则主要由尾部的感觉神经元构成。如果学习激活了介导路,那么突触强度的变化属于同突触变化,习惯化就是这种情况。如果学习激活了调制回路,那么突触强度的变化属于异突触变化,敏感化就是这种情况。
后来我们发现,经典条件反射同时包含同突触变化和异突触变化。实际上,同突触变化和异突触变化都是突触可塑性的基本形式,它们可以组合起来构成更加复杂形式,就好比我们用字母表中的字母组合成不同的词那样。
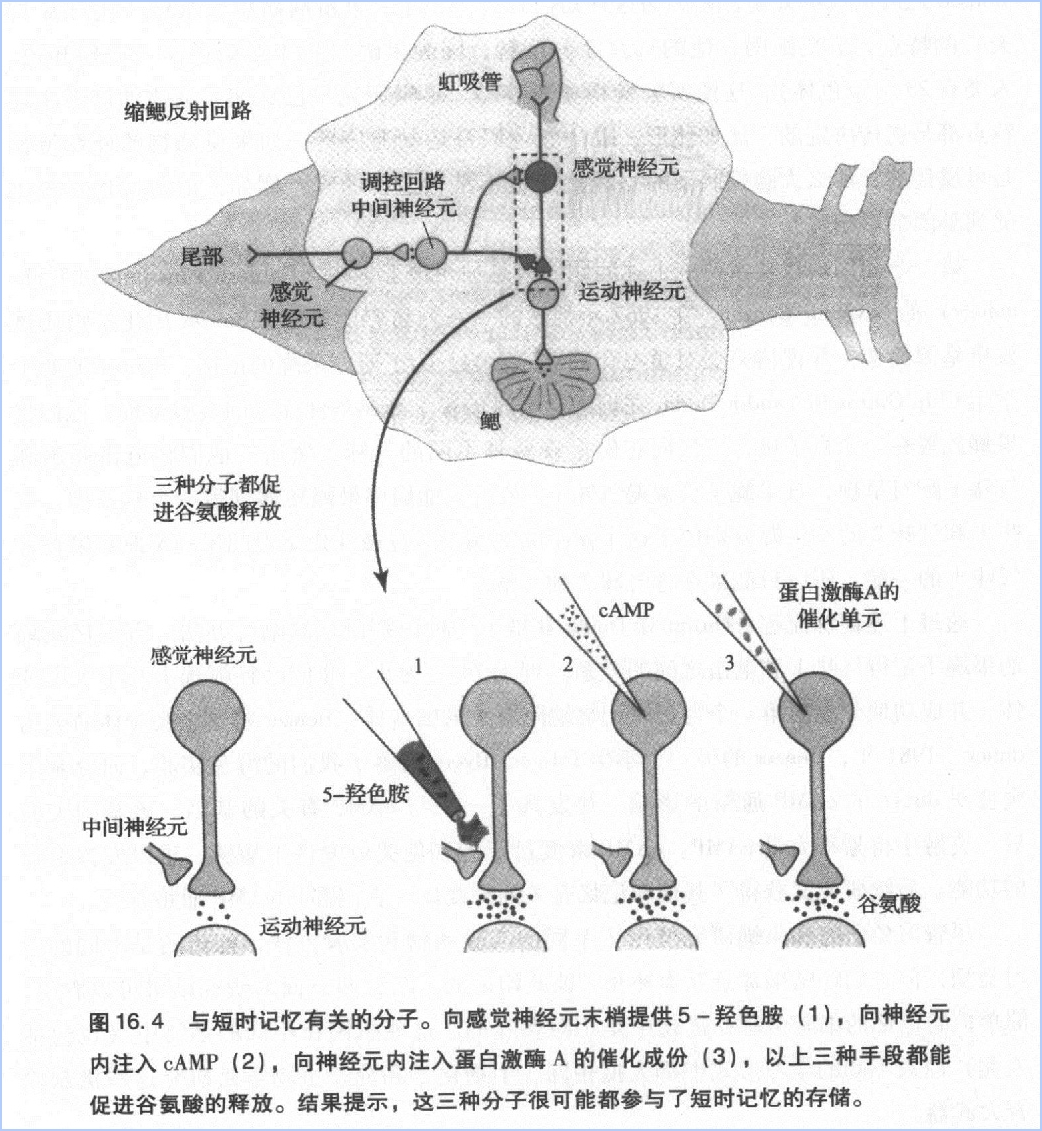
我开始理解为什么动物大脑里有如此丰富的化学物质,因为化学传导具有传导无法比拟的重要优势:它能够支持更多的学习形式和记忆存储形式。缩鳃反射运动神经元和感觉神经元之间的突触参与了多种形式的学与过程,显然,它的可塑性要比那习无关的神经突触更高。我们的研究证明,在能够被学习调控的神经回路中,只需少量训练,就可以让突触强度发生巨大且持久的改变。
记忆的形成分为不同的阶段,这是记忆的基本特钲之一。短时记忆可持续数分钟,长时记忆时持续数天或更长的时间。行为研究显示,短时记忆能发展为长时记忆。当然,反复练习是必需的,正所谓“熟能生巧”嘛。
对于艾宾浩斯和詹姆士的追随者来说,长时记忆的研究前景已逐渐明朗:下一步,应阐明长时记忆的建立过程。如今我们把这个过程称为固化。记忆若要持久,大脑必须对记忆内容进行深入彻底的加工。与此同时大脑必须对记忆内容投入更多的注意资源,还要将新信息与原有的知识网络联系起来。
他们的研究结果提示,练习后的1-2小时内,记忆内容还停留在短时记忆中,或者只是进入到长时记忆的早期阶段,因此很容易被干扰换句话说,长时记忆的化需要1-2个小时的时间。固化完成后,记忆就不容易被扰了。
综上所述,记忆储存的两阶段观点得到∫多方面证据的支持:记忆包括一个持续数分钟的短时记忆和一个持续数天以上的长时记忆,前者通过固化过程转化为后者,固化过程需要合成新的蛋白质。
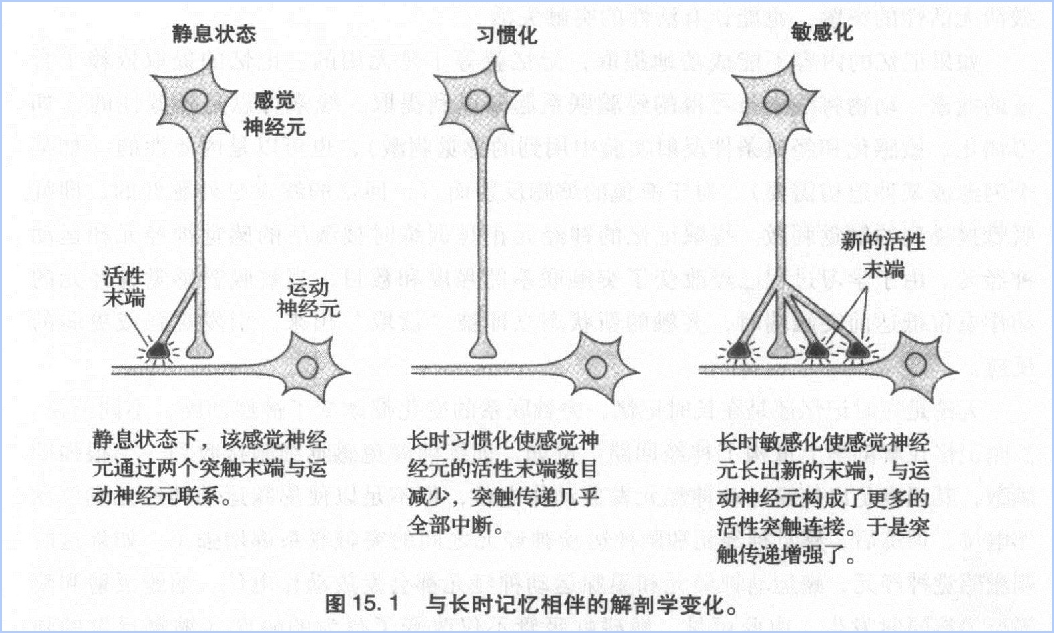
解剖学上的变化表现在几个方面。Bailey和Chen发现,一个感觉神经元拥有大约1300个前突触端,与大约25个靶细胞发生联系,其中包括运动神经元、兴奋性中间神经元以及抑制性神经元。在这1300个前突触端中,只有40%左右有活性,也只有它们能够释放神经递质。其余的前突触端则处于静息状态。发生长时敏感化时,前突触端的数目增至平时的两倍左右(从1300个到2700个),活性突触的比例也由40%增至60%。此外,运动神经元也长出新的突触末端。当记忆消失时,前突触端的数目由2700个降低至1500个左右,或者降到比平时稍高一点的水平。比平时多出来的这部分前突触端,也许能够解释艾宾浩斯发现的那个现象:第二次学习要比第一次学习容易。另一方面,发生长时习惯化时,前突触端的数目从1300个降至850个左右,活性突触的数目也由500个降至100个,这意味着突触传递几乎完全停止(图15.1)。
如果记忆的内容不能成功地提取,记忆就等于是无用的。记忆的提取依赖于合适的线索,动物将线索与习得的经验联系起来以利提取。线索可以是外源性的(如习惯化、敏感化和经典条件反射实验中用到的感觉刺激),也可以是内源性的(如某个闪念或某种迫切需要)。对于海兔的缩鳃反射而方,回忆的线索是外源性的,即虹吸管接受到的触觉刺激。提取记忆的神经元正是训练时被激活的感觉神经元和运动神经元。由于学习过程已经改变了突触联系的强度和数目,当虹吸管感觉神经元的动作电位抵达前突触端时,突触的新状态立即被“读取”出来,引发更强或更弱的反应。
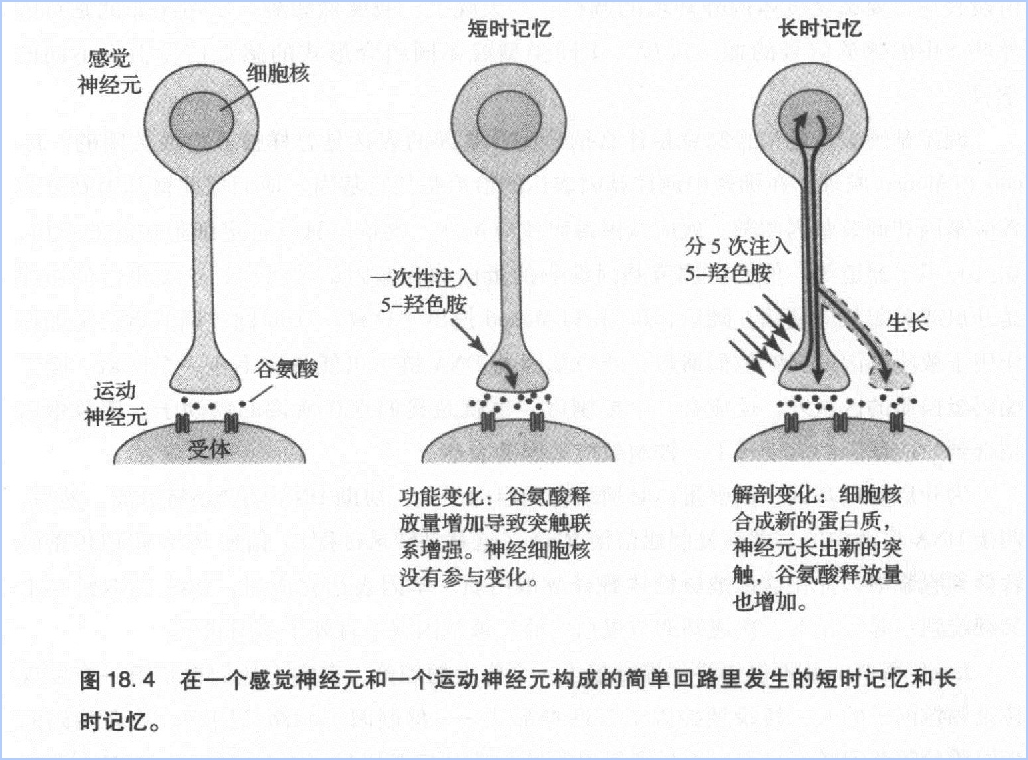
我们已经知道,记忆是神经回路内突触变化的结果:短时记忆是突触功能变化的结果,长时记忆还需要结构的变化。
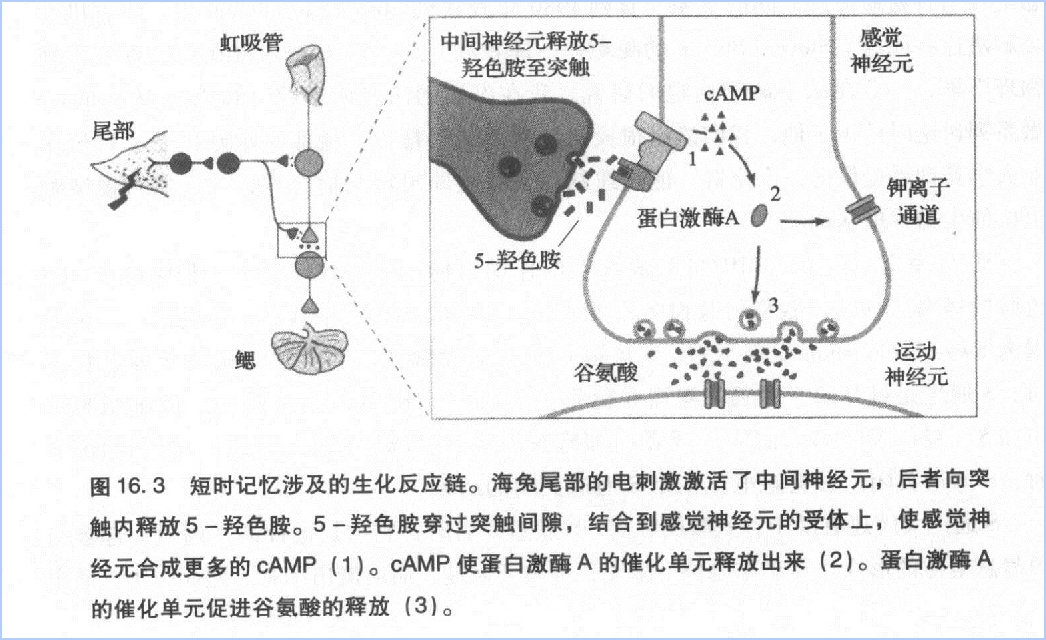
感觉细胞通过增加谷氨酸释放量的方式增强了突触后电位,使突触后细胞更容易产生动作电位,起到易化缩鳃反射的作用。
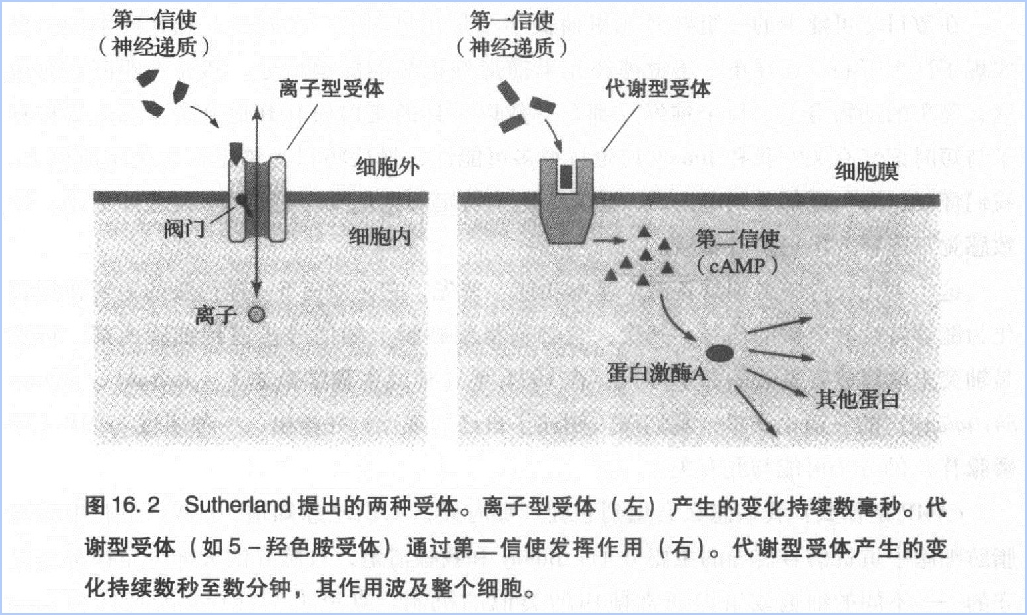
与离子型受体相比,代谢型受体的作用更强,作用范围更广泛,作用时间更持久。一般情况下,离子型受体的作用只能持续几毫秒,而代谢型受体的作用能持续几秒到几分钟,后者是前者的1000到10000倍。
Sutherland将与代谢型受体胞外端结合的化学信使称为第一信使,把细胞内的cAMP称为第二信使。他认为,第二信使将第一信使携带的信号从细胞表面转移至细胞内部,并在整个细胞范围内启动一系列反应(图16.2)。第二信使理论启发了我们,也许代谢型受体和cAMP就是慢突触电位和谷氨酸释放之间的中间环节。
蛋白激酶的作用是给蛋白加上一个磷酸基,也就是人们常说的磷酸化过程。磷酸化影响蛋白质的活性,磷酸化后,某些蛋白被激活,某些则失活。Krebs发现,磷酸化过程是可逆的,它完全可以作为分子开关,控制蛋白的活性。
接着,Krebs又深人探讨了这个分子开关工作机理。他发现,蛋白激酶A含有4个单元—两个调控单元和两个催化单元。磷酸化是由催化单元完成的。调控单元不具备催化功能,它只是“坐”在催化单元上,抑制催化单元的活动。调控单元上具有cAMP结合位点。当胞浆内的cAMP浓度增加时,调控单元上结合了过多的cAMP分子。在大量cAMP分子的作用下,调控单元的形状发生改变,从催化单元上滑落,它对催化单元的抑制也因此而解除。
正如分子遗传学家 Francois Jacob指出的那样,进化不是一名设计师,它不会发明整套全新的机制来解决新问题。进化只是一名修补匠,它反反复复地使用旧有的基因解决新问题。进化的工作方式是,从众多随机突变中筛选出有益的突变,使蛋白质发生轻度改变或者使细胞内蛋白质的分布发生变化。实际上,有益突变在全部突变中所占比例甚小,大多数基因突变都是中性、甚至是有害的。为数不多的有益突变增强了生物的生存和繁殖能力,使持有该突变基因的生物在自然选择中占据优势。Jacob曾这样写道:
人们常常把自然选择比作工程师。其实这个比喻一点也不贴切。首先工程师根据预先制订的计划行事。其次,工程师在设计新机器的时候不一定非要以老机器为基础。比方说,灯泡不是从蜡烛发展而来的,喷气式引擎也不是从内燃机发展而来的……最后,工程师在新技术的帮助下重新制造了一个物体,其水准达到了当时的完美标准—至少优秀的工程师都是这样的。
进化的工作方式与工程师不同,它从不照着草稿制造东西。身边的东西就是它的材料,有时它稍微调整一下现有的系统,使其具有新的功能;有时它则是将几个系统组织到一起,构成一个更加复杂的系统。如果非要把进化比喻成什么,也许修补匠是最贴切的选择,也就是我们法语里的bricolage。工程师使用的原材料和工具都是事先规定好的,修补匠就不一样了,他们使用的都是一些零七八碎的东西………他们用纸板、绳子、木头片或者金属片等身边常有的东西制作出能用的物体。修补匠们并不刻意收集某些材料,他们能运用零散的材料创造出意想不到的物体。如果他们用旧轮胎做出一把扇子或者用破桌子做成一把阳伞,你完全不用感到意外。
生物通过对现有的分子进行简单修饰或调整分子间的相互作用来创造新功能。长期以来,人类的高级思维活动一直被视为自然界独一无二的功能,因此,许多早期神经学家希望在人类的大脑灰质中发现独特的蛋白质。然而,科学证明,人脑中几乎没有独特的蛋白质或者信号传递系统。脑组织含有的蛋白与身体其他组织含有的蛋白大致相同,几乎所有的大脑蛋白都能在其他组织中找到,这些蛋白质的功能也极为类似即使对于大脑的特殊功能也是如此。例如,作为神经递质受体的蛋白质就不是脑组织所特有的。这就像搭积木一样,所有生物的思维和记忆都是由相同的结构单元构成的。
9年后,在1953年4月25日Watson和Francis Crick在《自然》上公布了DNA的结构模型。在生物学家 Rosalind franklin和Maurice Wilkins拍摄的X-射线衍射图的帮助下,Watson和Crick推断出,DNA由两条长链构成,两条链相互缠绕,形成螺旋型的结构。他们知道,DNA的双螺旋结构是由4种反复出现的核苷酸构成,它们含有不同的碱基,分别是腺嘌呤(A)、鸟嘌呤(G)、胸腺嘧啶(T)和胞嘧啶(C)。Watson和Crick设想,基因中编码遗传信息的元素可能就是这4种核苷酸。后来他们发现,DNA的双链是互补的,一种碱基只能特异性地与补链上的另种碱基结合,形成碱基对。具体地讲,腺嘌呤只能与互补链上的胸腺嘧啶结合,而鸟嘌呤只能与补链上的胞嘧啶结合形成碱基对。一串碱基对牢牢地将两条DNA链联系在一起。毋庸置疑,Watson和Crick提出的DNA双螺旋模型和他们的惊人发现对当代分子生物学起到了多么深远的影响。
Watson和Crick的双螺旋模型向我们展示了DNA的复制方式。复制时,双链DNA要先解开成为两股链,然后以每条链为模板合成新链。我们称原来的两股DNA链为父链,新合成的两股DNA链为子链。由于碱基配对的方式是固定的,那么当父链的碱基序列已知时,我们就能推出子链的碱基序列:A对应T,G对应C。随后,子链也可作为模板合成新链。细胞分裂时,DNA以这样的方式扩增,获得多个自身拷贝,并将这些拷贝分配到各个子代细胞中。这种复制形式存在于所有组织细胞中,其中也包括精子和卵子。我们知道,精子和卵子是生殖细胞,因此,这种复制方式确保了生物体自我复制的能力。
Watson和Crick还进一步阐述了蛋白质合成的机制。他们推断,DNA的碱基序列中携带着蛋白质的密码,指导着蛋白质的合成。复制时,基因携带的密码通过合成互补链的方式被“读出”。然而,如后来的研究显示的那样,在蛋白质合成过程中传递遗传密码的分子并不是DNA,而是一种叫做信使RNA(核糖核苷酸)的分子。像DNA样,信使RNA也是由4种核苷酸组成的。其中三种核甘酸的碱基与DNA完全一样腺嘌嘌呤和胞嘧啶,但胸腺嘧啶被尿嘧啶取代。DNA的两股链彼此分开后,只有其中一股被转录成为信使RNA。细胞再根据信使RNA的碱基序列合成蛋白质。就这样,Watson和Crick阐明子生物学的中心法则:DNA制造RNA,RNA制造蛋白质。
20世纪70年代末,哈佛大学的Walter Gilbert和剑桥大学的Frederick Sanger发明了一种快速检测DNA碱基序列的新技术。这是一项很了不起的发明。从此,人们就能相对容易地读出某段DNA的碱基序列,并推测出这段基因编码的蛋白质。借助这项新技术,科学家们观察到,相同的DNA序列出现在不同的基因里,为各种不同的蛋白质编码完全一样或相似的区域。人们将这些可识别的区域称为结构域。相同的结构域行使相同的功能,无论它们出现在什么样的蛋白质中。现在,科学家们已经能够根据基因的碱基序列大致推测出蛋白质的某些重要蛋白质是不是激酶、离子通道或者受体。此外,通过比较蛋白质的氨基酸序列,还能推断出两种蛋白质的相似程度。
人们比较了很多物种的基因序列后,一幅描绘细胞工作机理和信号传递机制的蓝图逐渐浮现出来,为更好地理解生命过程构建起理论框架。尤为重要的是,上述研究再次显示,不同的细胞(从大处看则是不同的生物)由相同的物质构成。例如所有的多核生物都有合成cAMP的酶,都有激酶和离子通道。实际上,线虫、果蝇和海兔拥有人类基因组中约半的基因。鼠与人的基因相似度为90%,而高等猿类与人的基因相似度达到了98%。
调控基因表达的信号究竟是什么信号呢?基因的表达是怎样被开启或关闭的?Jacob和Monod发现,在细菌中调控基因表达的恰恰是其它基因。他们据此将基因划分为效应基因和调节基因两种。效应基因编码效应蛋白,也就是具有特定细胞功能的蛋白如酶和离子通道等。他们将调节基因编码的蛋白称为基因调节蛋白,这类蛋白的功能是开启或关闭效应基因。随后,Jacob和Monod提出了这样个问题:调节蛋白是如何作用于效应基因的呢?他们猜想,效应基因的DNA链上可能具有不止一个区域,除编码蛋白质的区域外,还应有—一个控制区,也就是我们现在所说的启动子。调节蛋白结合到效应基因的启动子上,控制效应基因的表达。
为开启效应基因,调节蛋白必须结合到启动子上,协助DNA的两条链分离。然后,两条DNA单链中的一条被复制成信使RNA(这就是转录过程)。信使RNA将遗传密码转移到胞浆中,并在这里被核糖体翻译成蛋白质。基因表达完成后,DNA的双链马上又缠绕到一起。在下次遇到调节蛋白之前,该基因将一直处于关闭状态。
在 Jacob-Monod的模型里,环境中的信号激活调节蛋白的基因,新合成的调节蛋白作用于效应基因,开启效应基因的表达,合成效应蛋白。我和Goelet大胆地设想,在敏感化长时记忆中是不是也存在着类似的信号和类似的调节基因呢?大家都知道,反复训练对长时记忆的形成来说非常关键。我们怀疑,训练过程可能导致环境信号传入细胞核,通知细胞核激活调节基因,并通过调节蛋白开启效应基因的表达,促进新突触的合成。如果我们的假设是正确的,那么记忆的固化过程就应该是调节蛋白开启效应基因的过程。此外,我的假设能解释为什么学习时或学习后的短时间内阻断蛋白合成可以阻止新突触的生长和长时记忆的形成。我们推测,阻断蛋白合成实际上是阻断了调节基因的表达。
反复训练使信号从突触转移至细胞核,而这个信号就是激酶。
尽管一直以来人们都认为基因是行为的主宰者,但我们的研究显示,基因是受制环境的。基因的表达受到外部事件的影响。环境刺激(如施加在动物尾部的电击)激活中间神经元,促进5-羟色胺的释放。5-羟色胺作用于感觉神经元,提高细胞内AMP的水平,促使蛋白激酶A和MAP激酶向细胞核内转移。在细胞核里,激酶激活了CREB,后者开启某些基因的表达,进而改变细胞的结构和功能。
1995年,Bartsch发现,细胞里存在着两种不同的CREB蛋白。与Jacob和Monod的理论相符,CREB-1激活基因表达,而CREB-2抑制基因表达。反复呈现的刺激促使蛋白激酶A和MAP激酶转移到细胞核里。在这里,蛋白激酶A激活CREB-1,而MAP激酶使CREB-2失活。看来突触联系的长时强化不仅需要开启某些基因,也需要关闭某些基因。
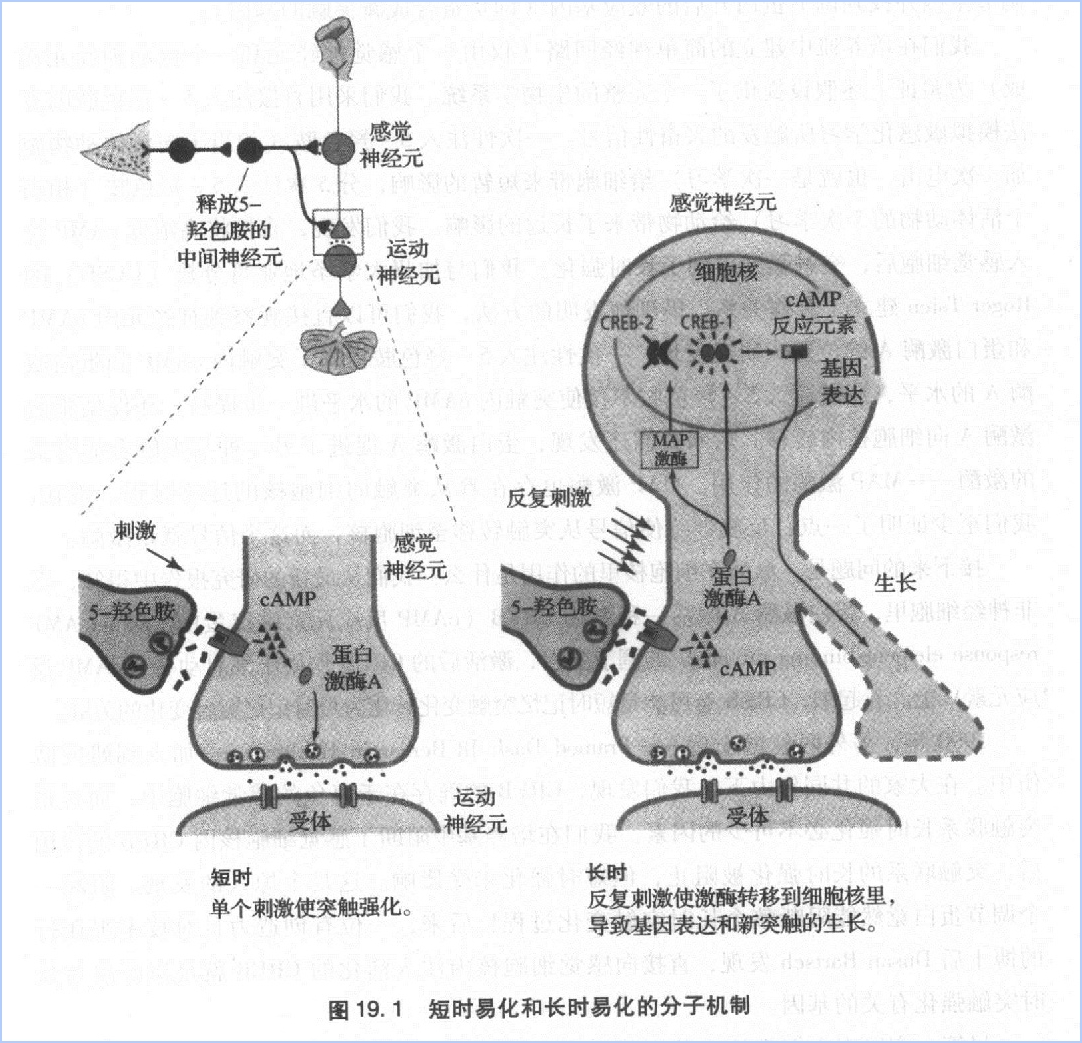
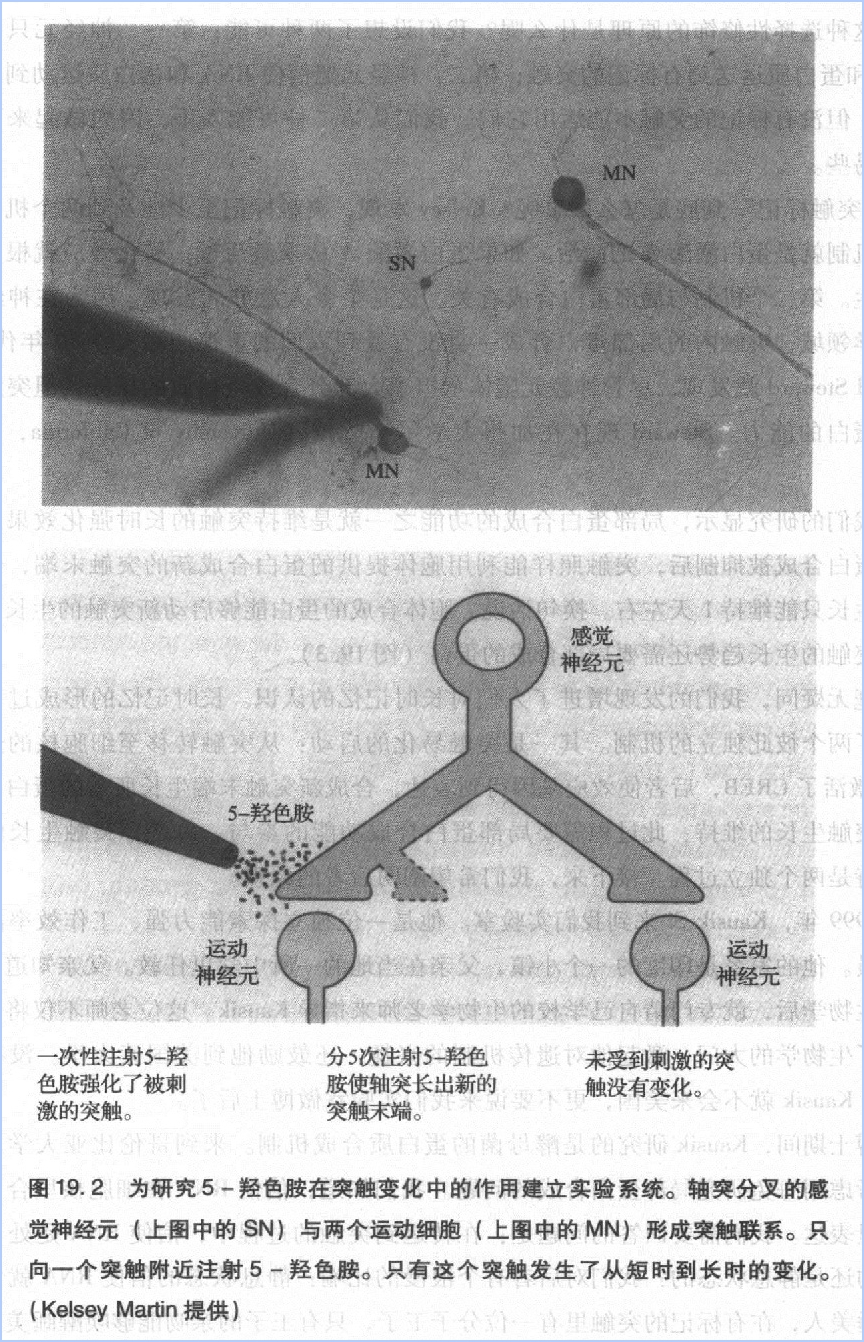
亳无疑问,我们的发现增进了人们对长时记忆的认识。长吋记忆的形成过程可能包含了两个彼此独立的机制。其一是突触易化的启动:从突触转移至细胞核的蛋白激酶A激活了CREB,后者使效应基因得到表达成新突触末端生长所需的蛋白质。其二是突触生长的维持:此过程需要局部蛋白合成功能的参与。总之,突触生长的启动和维持是两个独立过程。接下来,我们希望阐明后者的原理。
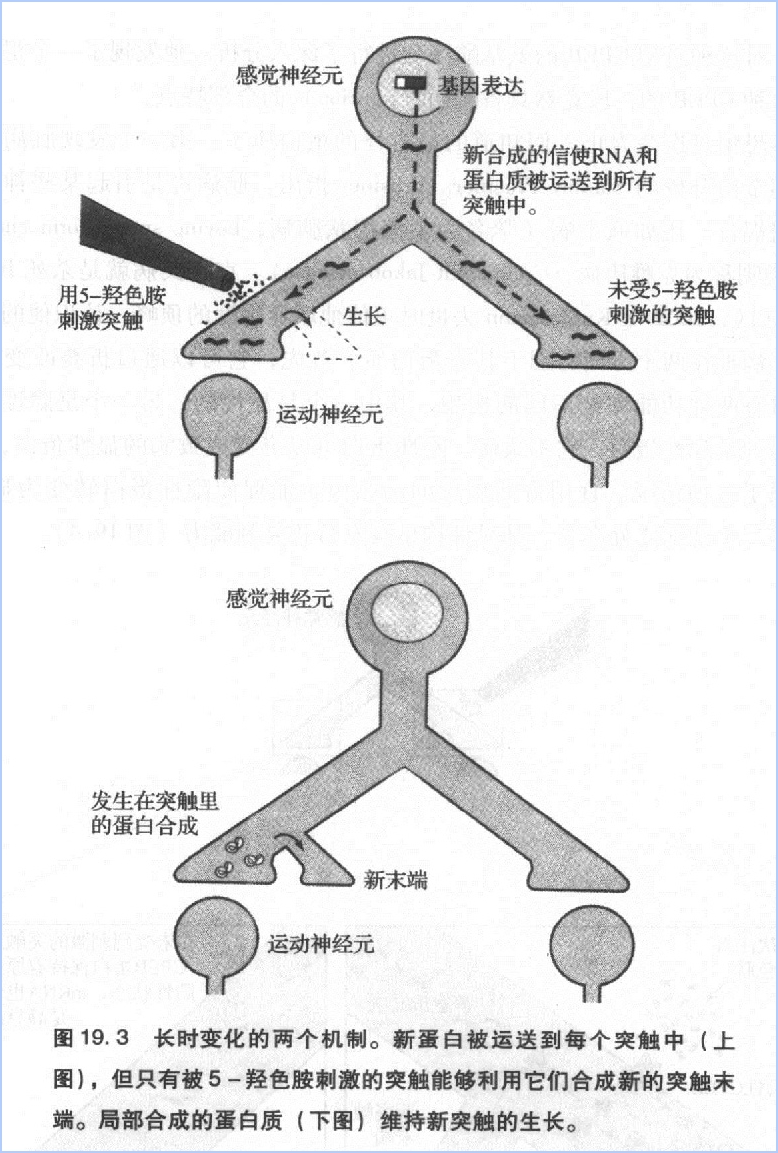
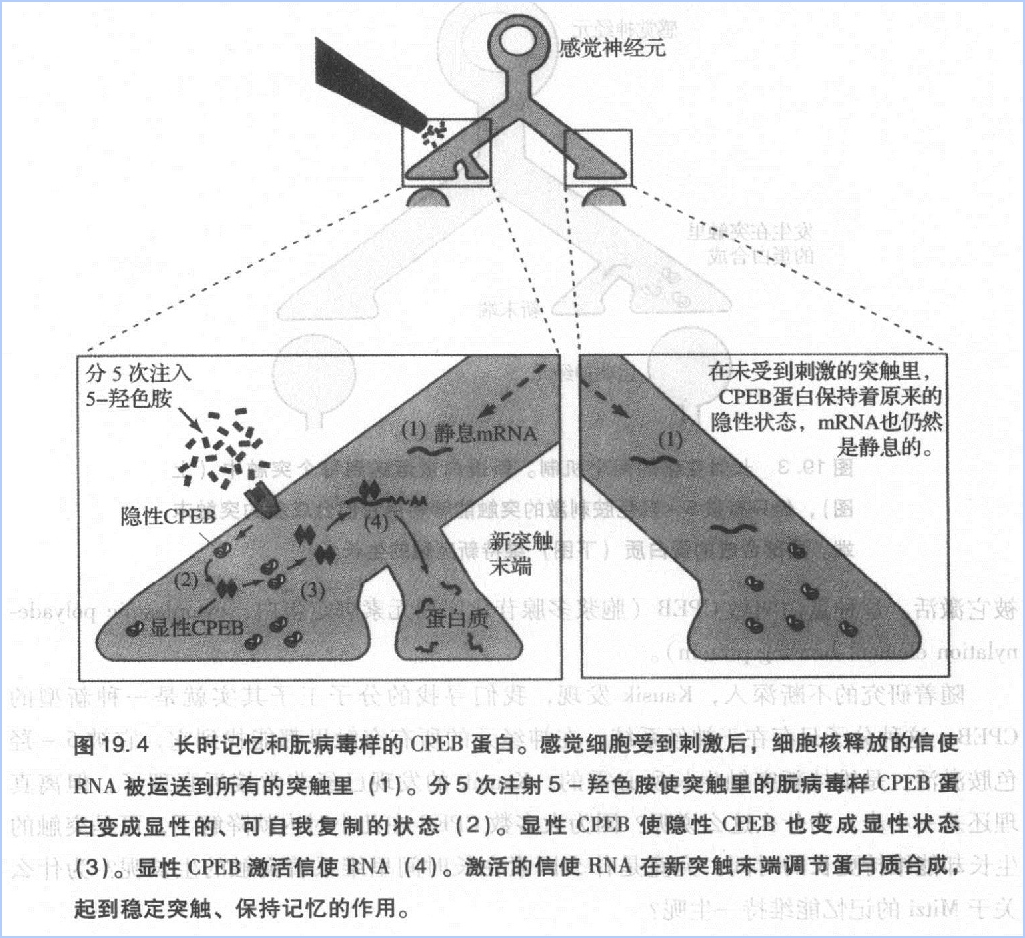
记得在2001年春天的一个下午,当我正透过办公室的窗户欣赏波光粼粼的哈得逊河时,Kausik走进来问我:“如果我告诉你,CPEB和朊病毒样蛋白很相像,你有什么感想?”
他不是在开玩笑吧?!但如果是真的,就不难理解为什么在蛋白质不断降解的情况下突触还能一直生长。显然,其有自我复制能力的分子可以无限期地存在于突触里,调节局部的蛋白质合成,维持新的突触生长。
朊病毒在酵母菌中研究得比较多。然而,在Kausik之前,没人知道这种蛋白质的功能。Kausik的发现不仅加深对学习和记忆的认识,还开辟了一块新的生物学研究领域。不久后,我们发现,在缩鳃反射的感觉神经元中控制CPEB构型变化的物质正是5-羟色胺(图19.4)。5-羟色胺使CPEB从无活性、无自我复制能力的形态转变为有活性我复制的形态,从而维持局部蛋白质合成过程。另外CPEB变成有自我复制能力的形态后,就不太容易再变回去了。
上述特点使CPEB非常适合于储存记忆,其自我复制能力使信息得以有针对性地、持久地保留在某个突触内,而不影响其它突触。Kausik很快就证明了这一点。
我们的研究证明了朊病毒与长时记忆的联系,扩宽了人们对大脑功能的认识。不仅如此,我们也为揭示朊病毒的本质做出了贡献。我们发现,普通的生理信号(如5-羟色胺)能改变CPEB的构型。其次,CPEB是第一个与生理功能有关的航病毒——具体到我们的研究,就是维持突触易化和记忆储存。在以往的研究中,有自我复制能力的朊病毒一般都是致病因子或是无活性的蛋白质。
下面我回顾一下本章的主要内容。我们的长时记忆研究得到三条基本原则,这些原则不只适用海兔,也问样适用于其它动物甚至人类。首先,长时记忆的形成需要开启某些基因。其次,不是所有的经验都能进入长时记忆。为开启长时记忆所需的基因,必须激活CREB-1蛋白,而起抑制作用的CREB-2蛋白必须失活。人们记不住所有的事情,也没人愿意记住所有的事情,所以编码抑制蛋白的基因肯定为经验设立了一个阈限,只有超过这个國限的经验才能进入到长时记忆中。这就是为什么我们只能记住重要事件的原因,生活中那些普普通通的事情往往都不会记得太久。排除这道阈限使短时记忆的内容顺利进入长时记忆。CREB-1激活的基因编码合成新突触所需的蛋白质。长时记忆需要开启某些其因的事实说明,基因不只是行为的决定者,它们还对环境(如学习)做出反应。
最后,形成长时记忆必须长出新的突触。如果看完这本书后你记住了某些内容,那么你的大脑一定又多了些突触。这种能力在进化过程中是相对保守的。比如,无论是人类还是较简单的动物,大脑的体表皮层图都受到感觉信息的调节而不断地发生变化。
第四卷
这些记忆中的景象…为什么在这么多年后都能保存得完好无损?莫非它们是由某些格外持久的东西构成的?——Virginia Woolf,《过去的圊画》(Sketch of the Past,1953)
我知道老鼠的空间记忆,就像海兔和果蝇的内隐记忆一样有两个组成部分:不需要蛋白质合成的短时记忆和需要蛋白质合成的长时记忆。现在我想要了解的是外显的短时和长时记忆是否有不同的突触和分了机制?海兔的短时记忆需要基于第二信使的短时突触改变。长时记忆则需要长时的突触改变,这种改变也需要基因表达的改变。
我和同事们观察了基因修饰小鼠的海马薄片,发现在任何一个通往海马的三大通道中,长时程增强都有两段与海兔的长时程易化相似。一列电信号引起短暂的早期长时程增强只能持续1到3个小时,因此并不需要合成新的蛋白质。神经元对这些电信号的反应与Roger Nicoll的发现一致:突触后胞的NMDA受体被激活,使钙离子流进入突触后细胞。这时钙离子作为第二信使引起了长时程增强,促进了现有的AMPA受体对谷氨酸盐的反应和新AMPA受体插入突触后膜。连续序列的电刺激信号产生一个后期的长时程增强,可以持续不止一天。我们发现这种后期增强的特征跟海兔突触增强的长时程易化很相似,那时这方面的硏究还比较少。在海兔和老鼠身上,这种后期长时程增强受调控中间神经元(Modulatory intemeuron)的影响很大。但是,老鼠的那些神经元释放的神经递质是多巴胺,而非羟色胺(又称血清素)。5-羟色胺是哺乳动物人脑进行注意和学习强化时的常用神经递质。跟海兔的5-羟色胺的功能一样,多巴胺激活海马内一种增加cAMP含量的酶。在海马中,增加的cAMP大部分出现在突触,细胞中,而在海兔身上则出现在突触前感觉神经元中。cAMP可以辅助蛋白激酶A和其他蛋白激酶,导致名为CREB的蛋白质的激活,并开启效应基因。
在我们研究海兔记忆时惊人地发现,有一种抑制记忆的基因可以产生CREB-2蛋白质。阻滞海兔这种基因的表达不仅可以使长时程易化的强度增大,也可以增加与长时程易化的突触数量。在海马中,我们也发现阻滞类似的抑制记忆的基因,也可增加海马中长时程增强和空间记忆的强度。
尽管内隐记忆和外显记忆明显的差别,但是无脊椎动物经过数百刀年的进化其内隐记忆储存的某些机制也与脊椎动物外显记忆的储存机制相似。伟大的生理学家Eccles很早就建议我不要放弃用无脑的海蜗牛研究哺乳动物的脑机制,因为记忆的某些关键的分子机制在所有动物身上都是相似的。
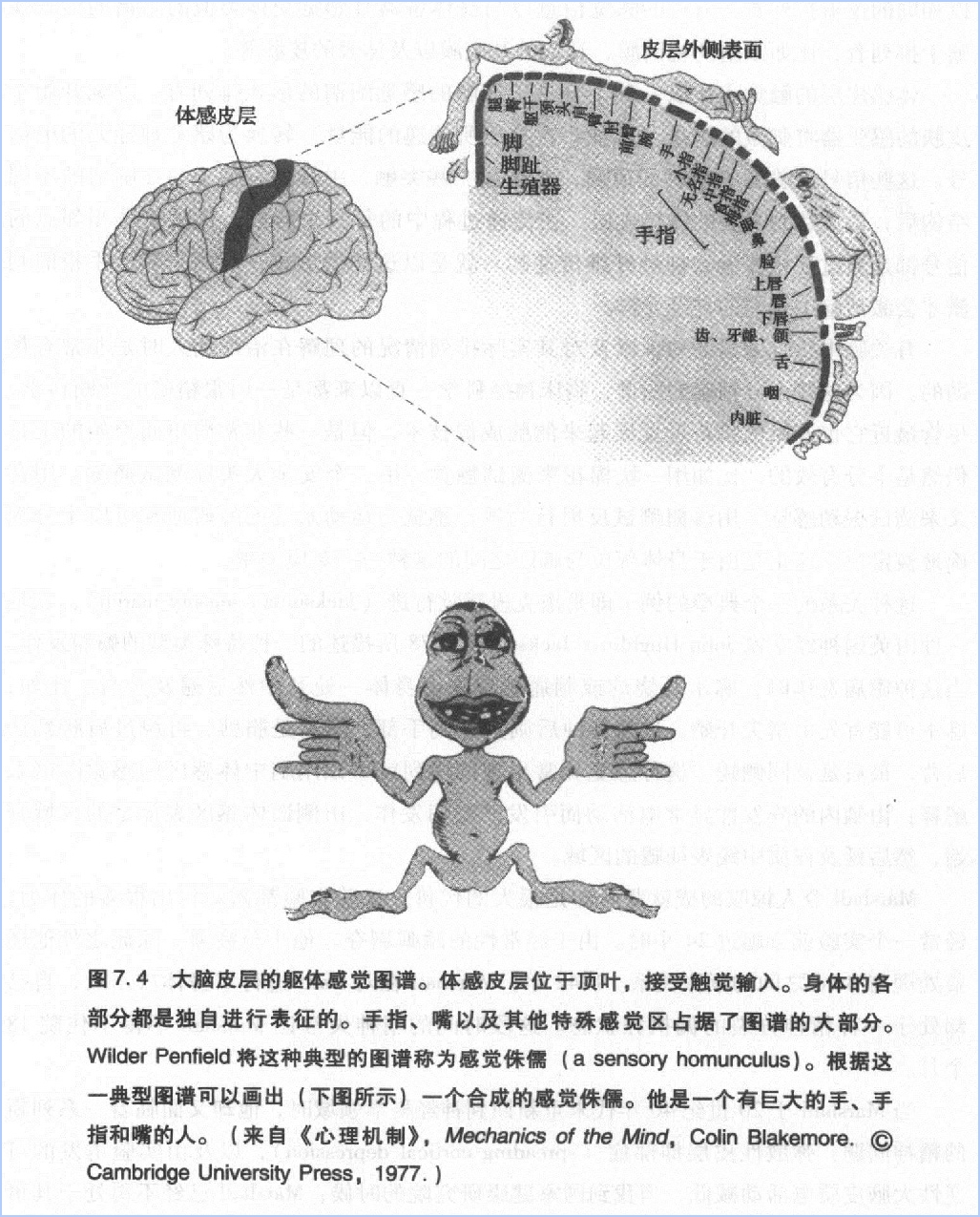
第一个进行感觉表征电生理研究的是我的导师Marshal,他主要研究触觉、视觉和听觉在脑皮层中的内部表征问题。1936年他发现躯体感觉皮层包含体表地图。与 PhilipBard和Clinton Woolsey一起,他在猴脑中发现了整个身体体表对应的表征;几年后Wilder penfield发现了人类体表对应的躯体感觉皮层。
这些生理学研究揭示了内部表征的两个原则:第一,人类和猴子身体的每一部分都以一个系统的方式表征在皮层中;第二,这些表征不是体表各部分在大脑中简单复制地形图,而是一个身体形式的生动变形,每一部分对应的比例关系不是依据真实尺寸的大小,而是依据其在感觉认知中的重要性。因此,指尖和嘴唇这些触觉敏感区占有一个与后背皮肤等不敏感区域完全不成比例的表征。Woolsey来发现在其他动物中也存在同样的变形:比如兔子的脸和嘴在大脑中的表征区域最大,因为这是它们探测环境时最基本的途径。因此,这些地图是随着经验而发生改变的。
在20世纪50年代早期,约翰·霍普金斯大学的Ⅴeamon mountcastle用单细胞记录法深入研究了内部表征问题,发现躯体感觉皮层中的单个神经元只对某一小块区域的皮肤信号反应,他称这个区域为这个神经元的“感受野”(receptive field)。例如,左脑的躯体感觉皮层的左手区细胞只对右手中指尖的刺激有反应,对其它区域的刺激不反应。
另外,Mountcastle注意到触觉可以进步分为多个亚型,例如,按压皮肤和用小刷子轻轻刷过皮肤引起的触觉分属于不同的亚型。他还发现每个不同的触觉亚型都有自己的脑内通道,它们彼此分离,在脑于和丘脑中传递并换站。Mountcastle发现躯体感觉皮层自上而下是以神经元功能柱的形式组织的,每一个功能柱指向一个触觉亚型和一块皮肤。因此一个功能柱中的神经元接收食指末端表面的触觉信息,而另一个功能柱中的神经元则接收食指的压力信息。另外,大脑皮层的结构有6层,穿过垂直的功能柱。每一层的功能柱有不同的联结模式。一些接受来自大脑中其它区域的信息,另些则发送信息到其它区域或其它的层。Mountcastle的工作揭示了感觉信息如何被解构,并发现每个触觉亚型是分别得到分析的,只有在信息加工的后期阶段才加以整合。他也提出,功能柱组成了皮层的基本信息加工模块,这个观点现在已经被广泛接受。
别的感觉形式也是如此组织的。目前我们对视觉通道的认识比别的感觉通道要多。Hubel和Wiesel发现视觉信息沿着从视网膜到大脑皮层的通路一点一点地传递,它也是得到精确地转换,先是被分解然后被重构,我们人类根本不会觉察到。
在20世纪50年代早期,Kofler在视网膜里用单细胞记录得到一个惊人的发现:那些细胞并不表达所有光信号,它们只表达明暗对比信号;最能够激活视网膜细胞的不是发散的光线而是光点。Hube和Wiee在下一步传递阶段的外侧膝状体核中也有相似的发现。并且,令人吃惊的是,一旦信号到达皮层,就需要发生转换,大部分皮层中的细胞对光点不反应,而是对具有线性特征的,比如直线、方块、正方形等比较敏感。
此外,每个皮层细胞都对线性刺激中的某个特殊朝向反应,有的对垂直朝向的线性刺激最敏感,有的则对水平朝向的敏感,也有的对有一定角度的线性刺激反应最强烈。Hubel和Wiesel接着发现,视觉系统像躯体感觉系统一样,具有相似属性的细胞(对相同朝向敏感的细胞)以功能柱的形式组合在一起。
这项研究非常了不起。作为一项脑科学的贡献,它是我们理解大脑皮层组织的重要基础。大脑皮层研究是Cajal在20世纪之交开始的,他发现个体神经元之间的联系是很精确的。Mountcastle、 Hubel和Wiesel发现,这些联系的模式有着重要的功能意义它们在皮层通路和皮层中过滤并转换感觉信息。皮层以功能层和模块的形式组织,这种组织结构可以随着经验发生改变。
视觉系统后续的研究发现,视觉其它方面,运动、深度、形状和颜色等都有同的通路通往大脑,在那儿整合为一致连贯的知觉。这种整合不是单一的分级排列,而是皮层的初级视觉区有两个平行的通路,在不同水平上互相影响通路是“what”通路,传递物体形状的信息。另一个是“ where”通路,传递物体的运动信息。这两个神经通路终结于负责更复杂加上的高级皮层区域。
那么,这些what和where通道中的有关动作、深度、颤色、形状等不同神经通路的信息如何整合为整的认知呢?问题与知觉经验的统一性有关,被称为绑定(binding问题:我们看个骑单车的男孩时,为什么不是看到一些没有画面的运动或者一个静止的面,而是看到个彩色的连贯的三维运动的景象。这可能是将几个担负不同功能的神经通路的信息临时绑定到一起,那这种绑定是怎样、在哪儿发生的?伦敦大学最出色的研究视知觉的学生 Semir zeki提出了这个问题:
一眼看上去,整合问题简单。逻辑上只是需要将来自不同视觉区域的信号结起,然后将结果给某个管理皮层区,这个管理区会综合各方面的信息提供给我最终的图像。但是大脑有自己的逻辑...如果所有的视觉区都向某个管理皮层区汇报,这个管理皮层区又向谁报?更直观地说,是什么负责“观看”这个管理区提供的视觉图像?这个问题不单单局限于视觉图像或视觉皮层。例如,什么在聆听听觉管理区提供的音乐,闻嗅觉管理皮层提供的气味?继续追寻这个庞大的想法是没有意义的,因为解剖发现:在视觉或者别的系统中,根本没有一个皮层区专门接受别的皮层区的汇报。因此,皮层必须运用不同的策略组织产生一个整合的觉图像。
当认知神经科学家观察实验动物的大脑时,可以看到哪个细胞在活动,并且了解这个大脑在加工什么。但是大脑是用什么策略读懂自己呢?这是意识属性的中心课题乃然是新的心理科学尚未解决的问题之一。
我们没有一个专门的空间感觉器官,然而空间的表征是十分精巧的认知感觉,涉及一个庞大的绑定问题。大脑必须综合几种不同的感觉输入形式最后得到一个完整的内部表征,决不单单依赖任何一种输入形式。大脑以不同形式在不同地方表征空间信但是每一种表征的特征却因目的而不问。例如,有的空间表征是动作指向表征,像转移视线到某个特定的位置、果蝈回避某种不喜欢的气味以及海兔的缩鳃反射等。对于这种行为,大脑一般是用自我中心式表征方法,例如在身体的哪个方向有与视野中央凹有关的光线、气味或触摸感来自哪里。相反,其它行为,像老鼠或者人的空间忆,就必须编码外部世界和有机体的位置关系,外部物体之间的位置关系,这时大脑采用的是世界中心表征。
选择性记忆是影响知觉、动作、记忆、意识经验的重要因素。当动物被大量的刺激包围时,就需要注意一个或几个,忽视或抑制其它的刺激。大脑加工感觉信息的容量比感受环境的受体数量少多了。注意像一个过滤器,只选择某一些事件进行深加工。很大程度上正是因为选择性注意,内部表征才不必复制整个外部世界,不必计划作出所有可能的行为反应。在每时每刻的经验中,我们集中注意在某些特定感觉信息上,将其它的排除在外。如果有人进来的时候,你的视线从书上转到他身上,你就不会注意到书上的字了,也不会注意到墙上的装饰和房间里的其他人。在回忆这一段场景的时候,你会更多地记得那个进来的人,而不是墙上的划痕。感觉器官的集中性是知觉的重要特征,正像詹姆士在《心理学原理》(The Principle of psychology,1890)中谈到的:“成千上万的事物……都罗列在我的感觉中,却从不曾进入我的经验中。为什么?因为我不感兴趣。我的经验是我同意参加……每个人都知道注意是什么。它以一种清晰活泼的形式占据了头脑。集中注意十分重要,它使我们能够在纷繁芜杂的环境中高效地处理特定的事情。”
大脑的注意机制什么?注意对编码重要的空间信息并且保持较长时间有什么帮助?我已经知道注意不是大脑的魔法,而是一个调节加工。国家健康研究院的Mickeyoldberg和Robert Wurtz发现,在视觉系统中,注意增强了神经元对刺激的反应。在注意相关的现象中有明显的以多巴胺为媒介的调节通路。产生多巴胺的细胞集中在中脑轴突投射到海马。我们发现阻滞多巴胺的活性,可以影响正在注意中的动物的空间地图。相反地,激活多巴胺受体能使没注意的动物空间地图变得稳定。中脑多巴胺能使神经元的轴突给很多区域发送信号,包括海马和前额皮层。
支持意识行为的前额皮层反馈信号给中脑,调节这些神经元的发放。我们的研究发现,其它与意识行为相关的脑区也能支持注意加工,因此,选择性注意对意识的同性很重要。
詹姆士在《心理学原理》中指出,注意至少包括两种形式:不随意注意和随意注意。不随意注意受到自动化的神经加工的支持,在内隐记忆中比较明显。在经典条件反射中,只有条件刺激比较突出时,动物才能学会将两个刺激联系起来。不随意注意可以被一些外部刺激的特征激活,像詹姆士说的“庞大的事物、明亮的事物、运动的事物或者血”。另一方面,随意注意(如注意自己正在行驶的道路和交通)是外显记忆的特征,它产生于内部需要,刺激特征不一定很突出明显。
詹姆士认为随意注意是一种有意识的加工,因此,它可能产生于大脑皮层。从还原论的观点看,两种注意都需要明显的生物学信号,比如调控神经网络功能和结构的神经递质。
我们对海兔、老鼠的分子学研究支持詹姆士两种注意的观点。两种注意的主要差别不是有没有显著特点的刺激出现,而是这种特点是否被意识感知到。因此当我驾车从我家到儿子家时,我意识到自己在专心考虑路线。但是当有一辆车突然出现在前方,我会不自觉地把脚放在刹车上。这些研究也表明,决定记忆是外显还是内隐的关键在于注意信号的使用方式。在这两种记忆中,从短时记忆转化为长时记忆都需要基因的激活,并且调节神经递质需要传递刺激的重要程度信号。收到信号后,基因就开启合成蛋白质,传送到所有的突触中。例如海兔是5-羟色胺诱发蛋白激酶A,老鼠是多巴胺诱发蛋白激酶A;不过这些显著信号对海兔来说,是进人内隐记忆,对老鼠来说,进入空间地图。
对内隐记忆来说,注意信号得以不自觉地收集,是一种自下而上的加工:在感觉神经元末梢被激活后,引起分子发生反应释放5-羟色胺。在空间记忆中,多巴胺自动出现,是自上而下的加工:大脑皮层激活细胞释放多巴胺,多巴胺调节海马的活动(图23.1)。
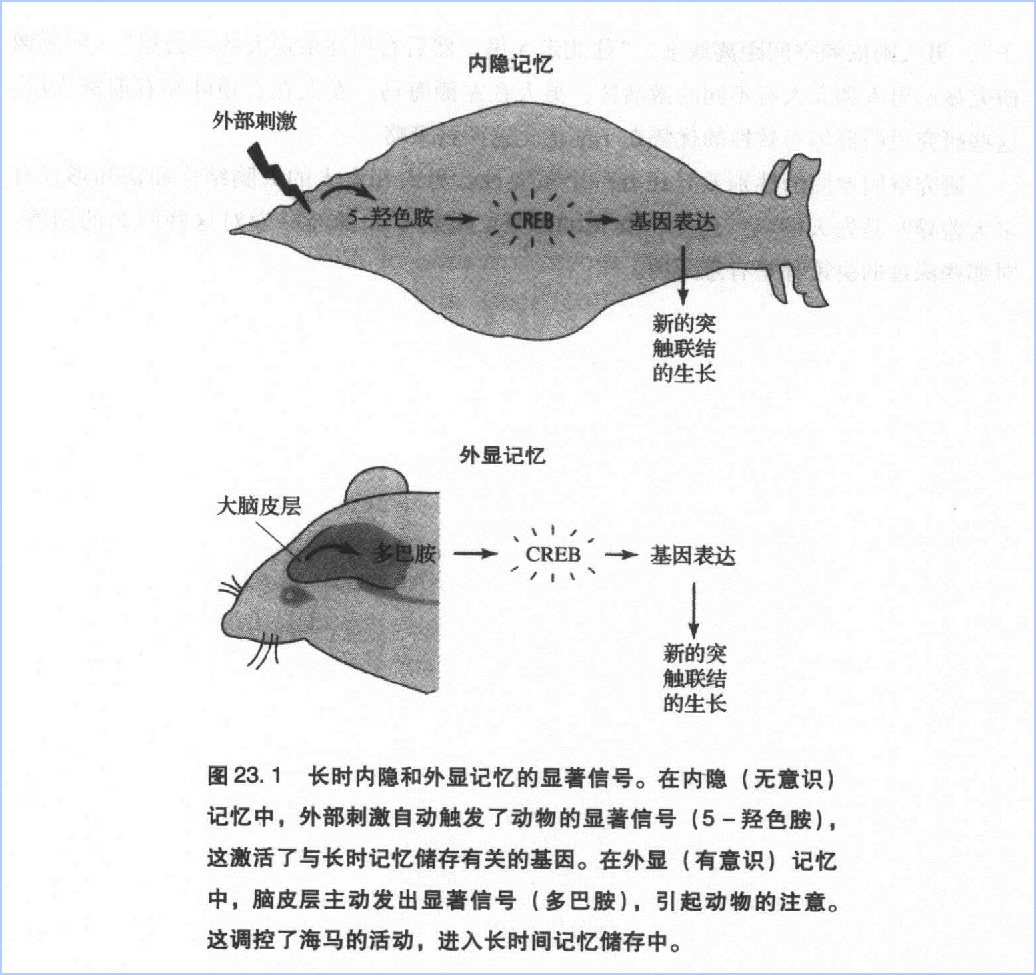
自上而下和自下而上的注意加工有相似的分子机制,同样地,在两种情况下记忆保持也有相似的机制。因此,与 Kausik Si在海兔身上的发现一样,老鼠海马至少也包含一种朊病毒蛋白(pion- like protein)。Martin Theis和我发现,海兔的5-羟色胺调节CPEB蛋白数量和状态的方式,与老鼠海马的多巴胺对CPEB-3蛋白质的调节是一致的。这时可以提出一个很有新意的假设:只有当动物的注意引起了海马释放多巴胺,多巴胺通过CPEB处于一种自我保持的状态时,空间地图才可能保持稳定。
第五卷
为找不到同样的动物模型,我们仍需要了解人性的很多方面。或许我们必须假设道德完全取决于上帝,如过人类是上帝約一个有机模型,那么研究我们自己可以使我们更多地了解上帝。——Sydney Brenner,诺贝尔获奖致辞(Nobel Lecture,2002)
早期阿尔茨海默症只有一些轻微的认知障碍,很难与良性衰老性遗忘症相鉴别。但是到了后期,记忆以及其他认知功能都开始大幅度衰退。这是因为突触联结的缺失和神经细胞的死亡。组织细胞的破坏是由脑细胞之间的一种异常—不溶性B-淀粉样变并堆积而成斑状物质造成的。
努力学习本身就可以增强常人的认知能力。
两种焦虑都可能引起病态。在威胁巨大又持久存在以致不能动弹时,本能焦虑就变得病态了;当习得性焦虑由非真实的威胁信息诱发,即中性刺激与本能焦虑发生关联时,也成了病态。因为焦虑症是最普遍的心理疾病,所以它吸引了我的兴趣。10%-30%的人一生中会受它折磨!
詹姆士反对这种观点,在1884年发表的那篇极有影响力的文章——《情绪是什么》(What is Emotion)中,他指出情绪的认知经验迟于情绪的生理表达。他认为,当我们遇到一个有潜在危险的环境,例如一只熊坐在路中间,对熊的凶残性评价,不会产生一个意识知觉的情绪状态。直到我们逃走,我们才感到害怕,我们身体先有本能反应然后才产生知觉,来解释这种身体反应。
基于这种观点,詹姆上和荷兰心理学家兰格(Carl Lange)指出,情感的意识体验发生在大脑皮层接受到身体生理反应的信号之后,或者说意识体验发生在某些无意识的生理变化(如血压、心率的升高或降低)之后。因此,当你看到火,你感到害怕是因为你的皮层刚刚收到身体心跳加速、膝盖发抖、流汘等。詹姆士写道:“我们感到难过是因为我们哭了,感到愤怒因为我们拍案而起,感到害怕因为我们发抖,而不是因为难过、愦怒、害怕而哭泣、拍案起和发抖。”根据这种观点,情绪是对中枢神经系统引起的机体变化的认知反应。我们的目常经验使我们确信机体信息引起了情感体验。
神经病学家 Antonio damasio修正了詹姆士-兰格的理论,他指出情绪体验是机体应的高级表征,可以是很稳定的,能持续相当长的时间。因此,人们对情绪产生达成了一致意见:首先是对某个刺激的无意识的、内隐的评估,随后引起了生理反应,最后出现了或持久或短暂的有意识体验。
通过对动物模型的研究,人们发现情绪的无意识部分是自主神经系统和负责调控的下丘脑的活动结果。在人类身上进行的情绪的意识研究发现,情绪意识是通过扣带回执行的大脑皮层的评价功能实现的。联结两个部分的就是位于皮层深处的杏仁核。杏仁核将情绪的意识体验和情绪的生理反应联结起来,特别是恐惧情绪。
对人类和啮齿类动物的研究表明储存无意识、内隐情绪记忆的神经系统与产生有意识记忆的神经系统不同。损伤了与恐惧情感有关的杏仁核,导致某种情绪刺激无法诱发相应的情绪反应;相反,损伤了与意识记忆有关的海马,就会导致不记得曾经出现过的刺激。因此,意识认知系统使我们可以选择采取哪种行为,无意识情绪评估系统将我们的选择局限在与当前情境有关的行为。这种观点的精华在于它使情绪研究与记忆储联系起来了。情绪记忆的无意识提取与内隐记忆储存有关,情绪记忆的有意识回忆则与外显记忆储存有关,需要海马的参与。
恐惧最主要的特征是它可以通过学习与中性刺激发生关联。一旦如此,中性刺激也可以诱发人们的长时情绪记忆。这种习得性恐惧是创伤后应激障碍、社交恐惧、广场恐惧(害怕空地)和舞台恐惧等的主要原因。在舞台恐惧及其它可以预期的恐惧中,未来事件(比如登台)与某些出错的预期(忘词)联系到一起,引起了恐惧。创伤后应激障碍往往是因为遭遇极端恐慌的事件比如生死搏斗、身体剧痛、抢劫,或者虐待。很显然当这种恐惧场景再次发生时,会令人们想起最初的创伤。这种障碍最主要的特征是创伤记忆多年来都很难磨灭,一旦出现危机就会勾起痛苦回忆,这是因为可怕的刺激激活了长时记忆。在创伤之后,负责情感加工的杏仁核可以在有机体的一生中都保持这种创伤这是如何发生的呢?
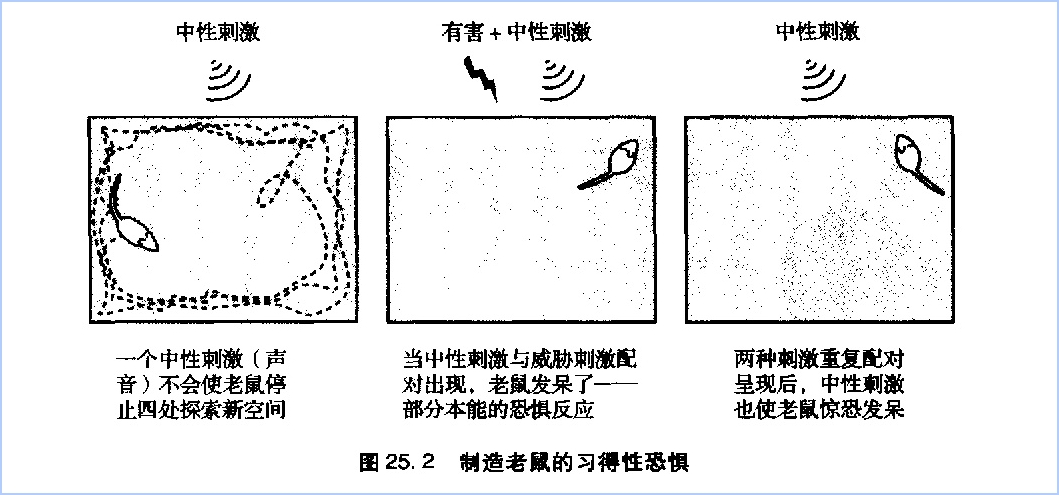
在纯音和脚部电击两种刺激配对呈现时,纯音信息(条件刺激)激活了听觉接受器官耳蜗的感觉神经元。这些感觉神经元轴突指向到负责听觉的丘脑神经元。丘脑的神经元形成两条通路通往杏仁核外侧核,这是杏仁核的最初的入口:直接通路从丘脑直接到外侧核,间接通路先到听觉皮层,然后到外侧核,从而包含了皮层内引起了意认知觉的区域(图25、3)。两个传递声音信息的通路终止于外侧核锥体神经元的突触联结。
来自非条件刺激——电击脚的疼痛信息激活了丘脑中另一条神经通路,该通路终止于一簇与痛觉加工有关的神经元。丘脑中的这些神经元也有直接和间接两条通路通往外侧核的锥体细胞。间接通路穿过了躯体感觉皮层。
两条分离通路的存在,一条穿过皮层,一条不穿过皮层,证明了恐惧信息的无意识评估先于有意识的皮层评估,跟詹姆士-兰格的理论是一致的。间接通路加工速度较慢,在我们通过间接通路意识到枪响之前,恐怖信息已经通过快速的直接通路引起了我们心动过速、流汗等反应。
除了作为条件刺激(声音)和非条件剌激(电击)传入信息的交汇点以外,杏仁核的外侧核也会评价威胁刺激并安排机体作出适应性反应,输出联系到两个重要的效应区:扣带回和下丘脑。下丘脑负责恐惧表达,作出“逃走还是抵抗”的反应(心跳加速、流汗、口干、肌肉紧张),扣带回负责恐惧评价。
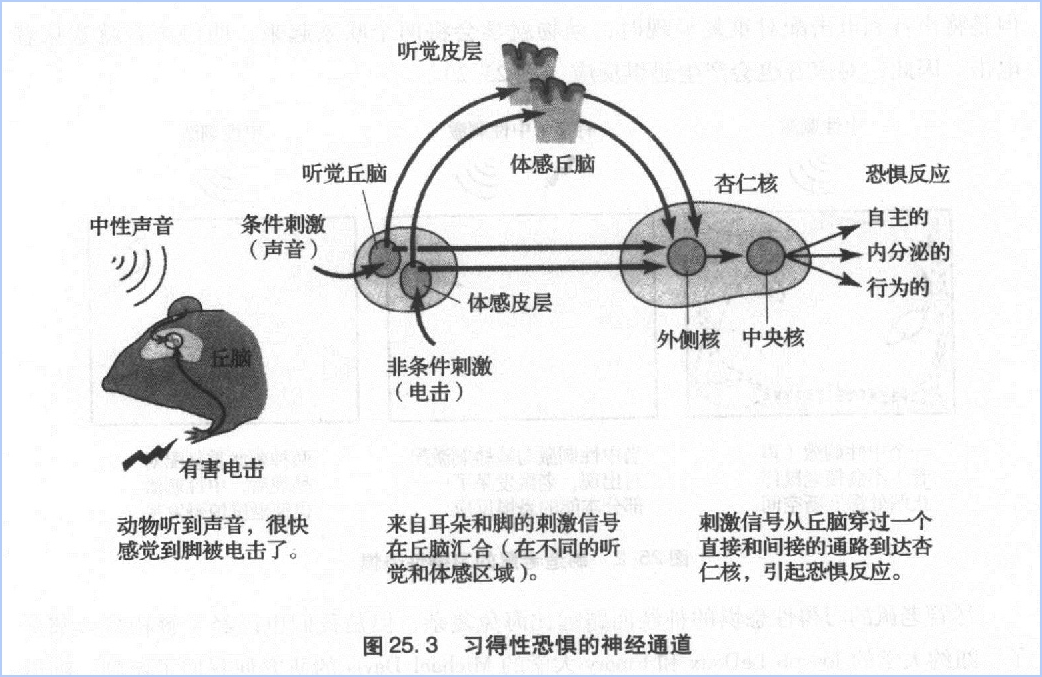
有了对习得性恐惧的解剖学和生埋学认识,我们更有信心寻找习得性恐惧的分子基础了。博土后 Gleb Shumyatsky和我开始寻找只在杏仁核的外侧核表达的基因。我们发现,外侧核的锥体细胞表达一种基因,编码一种称为胃泌素释放舦(gastrin-releaspeptide)的神经递质,以前它被看作是胃神经元的神经递质。现在我们发现,锥体细胞用这种肽作为谷氨酸以外的兴奋性递质,从它们的突触前端释放到靶细胞,而且些靶细胞是一群特殊胃泌素释放肽受体的抑制性中间神经元。像外侧核所有的抑制性中间神经元一样,它们的靶细胞释放GABA。这些靶细胞又与锥体细胞相连,一旦激活,就释放GABA抑制锥体细胞。
恐惧的反面是怎样的呢?安全感、自信、幸福是怎么回事?我不禁想到托尔斯泰的小说《安娜·卡列尼娜》(Anna karenin)的话:幸福的家庭都是相似的,不幸的家庭各有各的不幸。托尔斯泰认为焦虑和抑郁可以有很多形式,但是它们的反面——安全和幸福则具有相同的特征。
带着这种观点,Rogan和我开始研究一种幸福形式—安全感。我们的思路是这样的:既然在中性声音伴随着电击出现时,动物就会发现,声音预示着电击,那么如果声音和电击分别单独出现,即声音之后绝对不会出现电击,声音就应该预示着安全。实际上确实如此。在我们的实验中,老鼠慢慢明白声音是一个安全信号,当它听到这声音,就停止防御,走到盒子中央,仿佛那是它的地盘,不会表现出任何恐惧特征。我们观察这些接受安全感训练的老鼠的外侧核,结果发现种与时程增强相反的活动。声音在锥体细胞中诱发出一种长时程突触抑制,这表明向杏仁核传输的信号显著减少了(图25.4)。
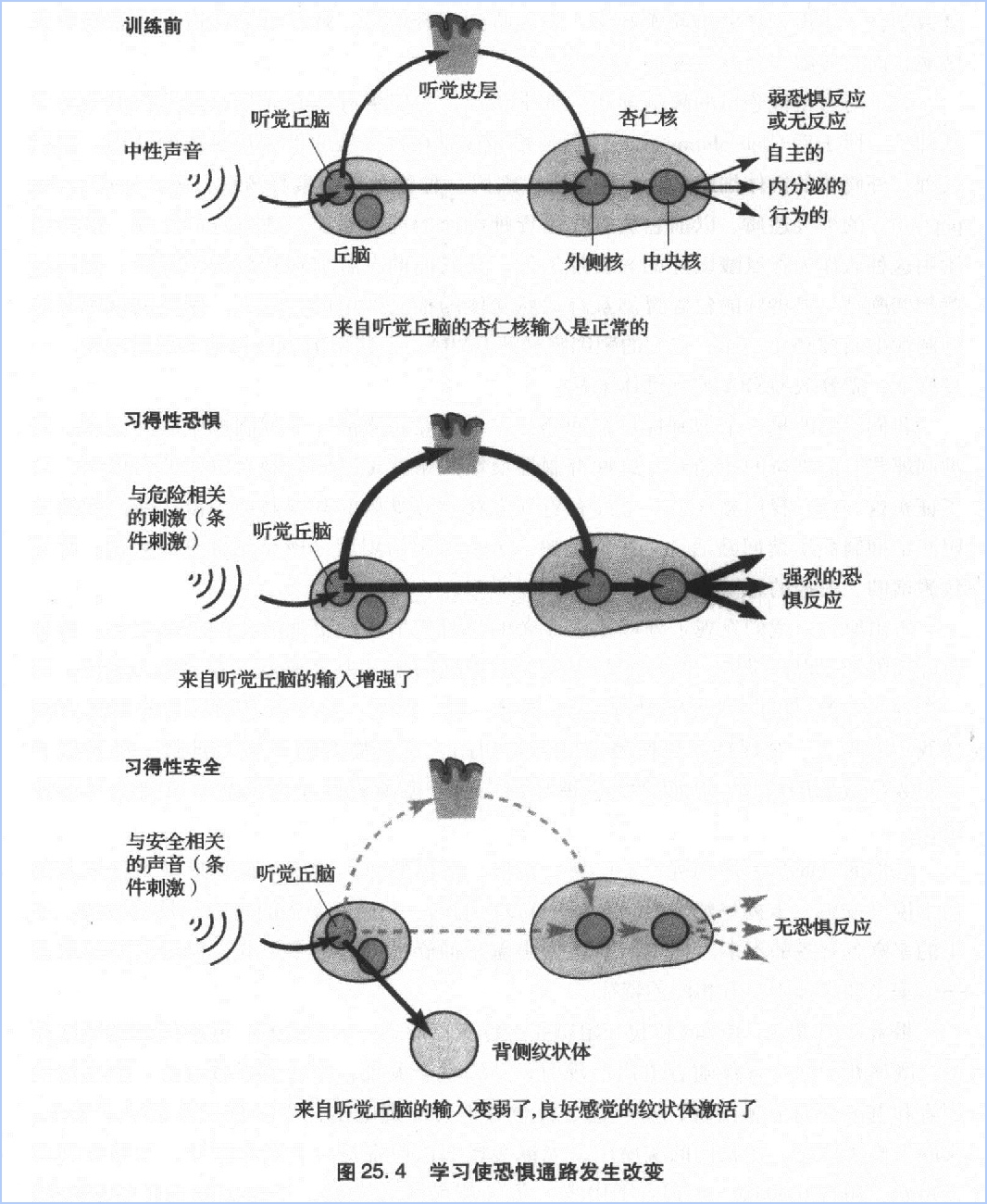
我们对习得性安全感的研究开辟了正性情绪和负性情绪研究的新视野。研究结果指向大脑深处负责正性情绪的第二系统。事实上,丘脑中对声调反应的神经元和杏仁核外侧核的神经元都发送联系到纹状体,传递满意和安全的信息。纹状体又与许多区域形成联系,包括抑制杏仁核活动的前额皮层等。所以可以推断,通过增强纹状体的信号,习得性安全不仅增强了安全感而且通过纹状沟与前额皮质的联系抑制了恐惧感。
精神分裂症主要有三种临床症状:阳性症状、阴性症状和认知症状。阳性症状临床中认为最少持续6个月,表现为怪异的行为和心理功能的紊乱,尤其是出现妄想和幻觉状态。这些都是患病期间最显著的症状,患者在此期间不能正确地认知现实世界,也就是说,患者不能真实面对他们的信念和知觉,无法将它们与自己身边世界实际发生的事情作对比。这种对现实世界感知能力欠缺的主要特点为妄想(一种明显违背事实的异常信念,并且此信念不会被能够明显让明其不合理的证据所改变)、幻觉(一种对没有实际发生的外部刺激产生的感知觉,比如听到别人对自己行为的评论)以及不合逻辑的思维(各种想法之间缺乏正常的联系,联系松散或者脱轨,严重时导致思维混乱语无伦次)。
精神分裂症的阴性症状表现为缺少正常的社交人际活动,伴随社交障碍、少言寡语以及情感淡漠等症状。认知症状包括注意力不集中和短时记忆/工作记忆的缺失。认知症状是慢性的、长期的甚至存在于症状缓解期,也是精神分裂症中最难以治疗的方面。
前额叶皮质与复杂行为的计划和执行有关一一这正是精神分裂症损害的脑功能,因此研究者们转向对精神分裂症患者前额叶皮层的探究。精神分裂症患者的脑成像显即使他们没有进行任何脑力活动,前额叶皮层的代谢活动也低于正常水平。当正常人遇到需要动用工作记忆的情况时,其前额叶区域的代谢功能剧烈增强;而同样情况下,精神分裂症患者的增加幅度则小得多。
组胺是一种细胞在应激反应时释放的类激素物质,能够引起血管扩张并降低血。他还指出,过量的组胺会引起些非常不利的麻醉副作用,诸如躁动、休克甚至突然死亡。
在Carlsson看来,这种过度活性与精神分裂症的所有症状——阳性症状、阴性症状和认知症状都有关联。Carlsson认为,海马和杏仁核与中脉的多巴胺通路中的过多的多巴胺可能导致了阳性症状;而中脑和皮层通路中的过量的多巴胺则可能导致阴性症状和认知症状的产生。研究者们很快发现,缓解精神分裂症状的药物主要针对一种特殊类型的多巴胺受体——D2受体。约輸霍普金斯大学的Solomon Snyder以及多伦多大学的 Philip seeman都发现了抗精神疾病药物的有效性和它们阻滯D2受体的能力之间的显著相关。同时人们也发现,这些药物仅仅对精神分裂的阳性症状有效,减轻了甚至消除了妄想、幻觉以及一些与阴性症状和认知症状联系不大的思维混乱表现。但是很难解释这些药物对于阴性症状和认知症状并无显著效果。
2004年,许多研究者发现,精神分裂症的遗传易感性在于,纹状体中D2受体的数量异乎寻常的多。纹状体与愉快体验有关。Simpson、 Kellendonk、 Polan和我决定探索此种遗传易感性对于精神分裂症认知方面所起的重要作用,所以我们用一个表达纹体区过量的D2受体的基因修饰小鼠,结果发现老鼠确实表现出Carlsson所假设的工作记忆缺失状态。
实验结果表明,在发育中过量的D2受体能引起老鼠大脑发生变化,并可以延续到成年。这些变化可能就是抗精神病药物对于精神分裂症的认知症状没有疗效的原因。在精神分裂症出现明显症状之前,纹状体中过量的D2受体可能已经在大脑其它部分多巴胺系统中形成一些不可逆的变化,影响了纹状体早期的发展。一旦这种情形发生额皮层的功能缺失便无法通过减少D2受体数量得以恢复。
治疗抑郁症的药物主要针对大脑中的两种神经递质,血液中的5-羟色胺和去甲肾上腺素。很显然,5-羟色胺与人们的情绪状态有直接的关系:血液中高浓度的5-羟色胺与健康的心态相关,低浓度的5-羟色胺同抑郁症相关。事实上,自杀的患者血液中5-羟色胺的含量最低。
耶鲁大学 Ronald duman和哥伦比亚大学 Rene Hen的研究工作促进了抑郁症的治疗进展。他们发现抗抑郁症药物增加了海马中的一块小区域——齿状回产生新细胞的能力。尽管绝大多数的干细胞没有分裂,但是这块位于海马中的小干细胞巢却分裂并分化成不同的神经细胞。经过大约两到三周时间,也就是抗抑郁症药物产生作用的时间,少量的细胞便进入齿状回的神经网络中。这些干细胞的功能到目前为止还不清楚。因此,Hen使用放射线破坏其海马齿状回的干细胞,结果发现,如果没有这干细胞,抗抑郁症药物便不能医治那些患抑郁症的老鼠。
贝克在临床中检验了这个方法,通过分析病人自身的经历、行为和后果来促使他们改变原有的负性观点。纠正病人的负面情绪,建立积极的思考模式,贝克发现病人有了惊人的改善,几个疗程后感觉好多了。于是贝克提出一个系统的抑郁症短期治疗方案,不是解决病人的无意识冲突,而是有意识地改变扭曲的认知。
贝克和同事们开始进行严格控制的临床实验,检验这种治疗方法同安慰剂和抗抑郁药物相比的有效性。结果发现在治疗轻中度抑郁症时,这种认知行为疗法与抗抑郁药一样有效,甚至在有些研究中,它能很好地防止复发。在后期的临床实验中,认知为疗法成功地治疗了焦虑症,特别是惊恐发作、创伤后精神紧张障碍、社交恐惧症、厌食症和强迫症。
贝克不仅带来一种新颖的精神疗法并得到实验验证,而且发展了抑郁症及其它精神障碍的程度与症状的评定量表,为精神疗法的研究带来新的科学研究方法。另外贝克和同事写了治疗手册。贝克为精神分析学带来一种至关重要的态度:重视实证数据以及心理治疗的有效性验证。
受贝克研究的影响,Gerald Klerman和Myrna Weissman建立∫第二种科学的短期精神疗法——人际心理治疗法。这种疗法致力于纠正病人的错误认识,通过与他人互动改变他们的思想。与认知行为疗法一样,严格的实验证实,人际心理治疗对轻度和中度的抑郁症有很好的疗效,并且这一疗法被编入教材。人际心理治疗对那些危机情境特别有效,比如失去配偶和孩子;认知疗法则专长于治疗慢性的心理障碍。同样,Peter sifneous和Habib davanloo也提出了第三种短时治疗方案—短程动力疗法(biedynamic therapy),专门针对病人的防御阻抗心理; Otto Kernberg发现一种针对移情的心理治疗。
与传统心理治疗不一样,这四种心理治疗都设法收集证据来支持其有效性。因此,他们完善了短期(甚至长期)疗法的有效性检验,开始推动学科向实证科学发展。
猴子和人类一样,有社会性发展的关键期。
婴儿的依恋系统真实反映了父母对婴儿情绪的敏感影响。父母的及时反应增强并巩固了婴儿的正性情绪状态,削弱了负性情绪状态。这些重复的经验编码到程序(内隐)记忆中,使婴儿感到安全。
生物学和精神分析学结合将增强精神病学在现代医学中的影响力,激励实证主精神分析学加入现代认知科学的学科联盟中。合并的目的是用激进还原论,带着理解脑的人文目标,推动基础生物学、精神病学和精神分析学。这也是脑科学的最终目的:通过对人类心理和经验构建进行全面深入的理解,就把我们自身和物理学、生物等自然科学研究紧密联系在一起。
为了得到更多意识的新发现,新的心理科学首先要将意识作为一种知觉状态或者明显的选择性注意进行操作定义。对人类来说,意识的核心是自我知觉,知道自己正在感知。因此意识是一种能力,不单单是体验快乐和痛苦,而是参与并根据这些体验作出反应,在我们当前的生活和整个人生中都是如此。意识注意使我们忽视无关的体验,集中在眼前的关键事件中,快乐或痛苦、天空的蓝色、 Vermeer画中冷色调的光线或沙滩的美景。
屏状核是注意的集中点,合并知觉的所有不同成分,是Crick生命中一系列重要思想中的最后一个。
恐惧面孔的无意识知觉激活了杏仁核的外侧基底核。与老鼠相同,杏仁核的这个区域接受大部分输入的感觉信息,是杏仁核与皮层相通的主要途径。对恐惧面孔的无意识知觉引起的外侧基底核激活程度取决于背景焦虑程度,背景焦虑越高,反应越强烈。低焦虑的人完全没有反应。相反地,对恐惧面孔的意识知觉,激活了杏仁核的背区域,包括中央核,并且激活程度与背景焦虑程度无关。杏仁核的中央核发送信息到负责唤醒和防御反应的植物神经系统的部分脑区。总的说来,无意识条件感知的威胁,对高焦虑人群有不成比例的影响,而意识条件下感知的威胁,则引起所有被试或战或逃的反应。
第六卷
真正热爱维也纳的人是靠虚构的记忆来生存的。他会怀着一种苦而甜蜜的怀旧伤痛,记着那些他从来不知道的事情……那虚幻中维也纳就是史上最壮啁的城市。——选自Orson Welles,“维也纳1968”(Vienna 1968)
在位于特尔斐(Delphi,希腊古都,因太阳神Aplo的神殿而著称—译者注)的阿波罗寺庙入口上方的石碑上曾刻着这样的座右铭:认识你自己。自从苏格拉底和柏拉图首先提出对人类精神本性的思考以来,一代又一代的思想家,从亚里士多德到笛卡尔,从埃斯库罗斯(Aeschylus,希腊的悲剧诗人)到斯特林堡(Strindberg,瑞典知名剧作家)和伯格曼(Ingmar Bergmar瑞典顶极电影导演),都认为了解自己的内心和行为是一件明智的事情。
Arvid、Paul和我,以及我们这一代的科学家已经试图用实证性的生物学语言来表达有关心理的抽象的哲学问题。指导我们工作的关鍵原则是明确心理是由脑控制的一系列操作定势(set of operations),脑是极度复杂的具有计算能力的器官,它建构我们对外部世界的知觉,支配我们的注意,控制我的行为。
我们三人通过研究单个神经细胞内部以及神经细胞之间的生物化学信号,与心理加工和心理障碍的关系,从而迈出了将心理加工与分子联系起来的第一步。我们已经发现脑内的神经网络不是固定的,神经递质分子可以调节和控制神经细胞之间的交流,而神经递质分子正是你们瑞典的分子药理学学院发现的。
展望未来,我们这一代的科学家相信认知神经生物学对于本世纪的意义就如基因生物学对20世纪的科学界一样重要。从更大的意义上来说,认知神经生物学研究不仅仅具有远大前景的科学探索,也是一种重要的人文主义追求。认知神经生物学在自然科学(有关自然世界)和人文学科(有关人类经历的意义)架起了一座桥梁。这种新的整合产生的见解不仅会增加我们对精神病和神经病学的了解,还将推动我们更深一步地认识我们人类自己。
事实上,甚至我们这一代都已经初步获得了更深层次地认识自我的生物学知识。我们知道,虽然那座右铭的宇迹已经不在特尔斐神庙前的石碑上,但是它们已被刻入我们的脑海中。这个座右铭已经被大脑中那些特定的分子加工过程保存在记忆中几个世纪了,而今天我们才刚刚开始注意到这些分子加工过程的存在,并开始了解它们具体的机制。
用几年时间去尝试新鲜的、原创性的事情,要比进行那些每个人都在做或是别人能和你做得一样好的常规实验来得更有价值。



